
Introduction
In the scientific field of paleoanthropology, and paleontology overall, being able to deduce how an organism moved is very useful. This is because it can show how an organism lived and can give insight into its evolutionary history. Most of the time, all paleontologists have are fossils, but this can be surprisingly helpful. For paleoanthropologists specifically, being able to do this is crucial, as it can help determine how a hominin is related to other species and therefore, its place in our family tree.
The field concerned with determining how an organism moved/moves is known as biomechanics. Biomechanics is the study of the structure and function of biological systems. When combined with paleontology, biomechanics can give great insight into how an extinct animal moved, pulling from an array of morphological characteristics. Biomechanic scientists look for and examine all these characteristics in fossils to get a good idea of how extinct organisms moved.
Animals move in all sorts of locomotor styles. Fish often move by slowly moving their tails back and forth to propel themselves through the water. Most land animals move quadrupedally (on four legs), and retain a pronograde posture. This means that their bodies are more horizontally positioned. Many animals that walk this way have sprawled out limbs, such as in lizards, whereas others have limbs tucked beneath them, such as in quadrupedal mammals like cats and dogs.
However, many animals walk bipedally (on two legs). Many theropod dinosaurs walked this way, including Tyrannosaurus rex and even modern birds. Some mammals walk this way too, such as kangaroos. This is the locomotor style of humans. There is one thing that makes us different from other bipedal animals however, and that is our posture. Humans retain an orthograde posture, rather than a pronograde posture. This means that our posture is upright/vertical. Some other primates, such as gibbons, possess an orthograde posture as well. Most of the time, gibbons move via suspensory brachiation, where they swing from branch to branch with their front arms. Gibbons also can move bipedally when need be, but it is not their main method of locomotion.
The first known hominin that possesses these locomotor styles is called Sahelanthropus tchadensis, which lived roughly 7 million years ago. Sahelanthropus, along with all the other fossil hominins, possess many different traits that allow for them to walk bipedally. These traits can be found from the head to the feet. Most are directly related to the animals’ locomotion, though some are indirectly related. Some even have other side effects not related to locomotion as a result. Lets go over all these traits from top to bottom:
The Skull:
The first trait is found in the skull. More specifically in the basicranium (the lower part of the skull). This trait has to do with the foramen magnum, the hole at the base of the skull where the spinal cord enters. In organisms that walk quadrupedally, their foramen magnum is more posteriorly positioned, meaning it’s towards the back of the skull. This allows for the spinal cord to enter more horizontally, giving the animal a more pronograde posture. The foramen magnum is also slightly angled. Depending on the locomtion of the animal, the hole will be angled back or forward slightly. The position tells you the posture and the angle tells you the locomotion.
In bipedal species with an orthograde posture, the foramen magnum is more anteriorly positioned, meaning it is closer to the front, and is angled forward slightly. The occipital condyles, small bony projections on each side of the foramen magnum are angled as well, allowing for articulation with the cervical vertebrae. Sahelanthropus possesses this trait, though to a lesser extent than modern humans.

The Rest of the Body:
The next trait is the curvature of the vertebrae. In most apes, their vertebral column forms a curved ‘c’ shape. This allows them to walk on four legs. In humans however, our vertebral column forms a more ‘s’ shape. This condition in humans is known as lumbar lordosis. This curvature in the lumbar (lowermost) vertebrae places the center of mass of the body directly over the hips and legs, and distributes weight throughout the body. This makes it easier to maintain an orthograde posture and bipedal locomotion.

This condition first appeared around 4 million years ago, with the genus Australopithecus.There are several specimens of Australopithecus that have well preserved vertebrae showing they had this human-like condition. A specimen of a juvenile Australopithecus afarensis dating to 3.3 million years ago shows a very human-like condition in the vertebrae. The specimen, DIK-1-1, was nicknamed “Selam” after the Amharic word for peace, or the “Dikika child”, after the place she was found. Selam was a very significant discovery, and showed lots about the evolution of human growth patterns, but most importantly, she had a fully articulated human-like vertebral column.

Though lumbar lordosis is beneficial when it comes to bipedalism, it doesn’t come without cost. Lumbar lordosis can cause severe pain and result in several medical problems, such as DMD (Duchenne muscular dystrophy) and walking difficulties.
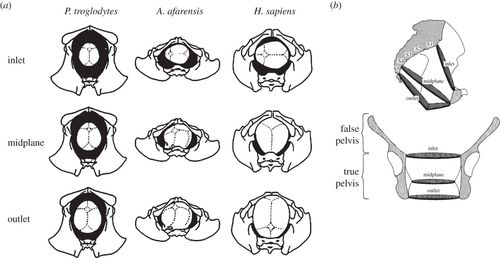
Lower down in the body, we find perhaps the most important trait: The orientation of the pelvis. In bipedal species, the iliac crests (the upper, wing-like part of the hip) are sagittally oriented. This means they are oriented into the midline of the body. This gives the pelvis a more bowl-like shape. This is further supported by a wider sacrum bone than in other apes. This allows us to balance upright without having to shift our weight forward, which would require us to be quadrupedal. It also plays a role in thermoregulation, as it lowers the surface area-to-mass ratio, allowing for more heat loss.
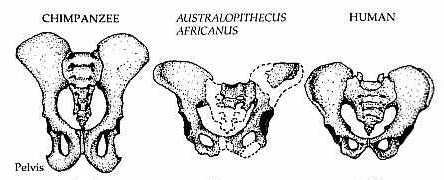
This evolutionary advantage comes with a trade off, an evolutionary compromise, however. With the narrow bowl-like pelvis, giving birth is much more difficult. In humans, the baby’s needs to rotate 3 times to fit through the birth canal, prolonging the average birth time to 14 hours, compared to other great apes, such as chimpanzees, which have an average birth time of 2 hours. Another reason why other apes have it so easy is because of the small heads of the babies being born. The ability to walk bipedally outweighed the need for short births, so we compromised one trait over another.
Below the pelvis, the femora (bones in the thigh) can also give a good clue of how a hominin walked. Organisms that walk bipedally possess a bycondylar angle, a slight angle in the femur that places the individual’s body mass beneath them, which is advantageous for this method of locomotion. Along with this, the epiphysis (the head of the femur) is thicker than in quadrupedal apes. This places the knee in a valgus (angled away from the midline) position. The condyles at the end of the femur are also thicker, allowing them to bear more weight while walking.
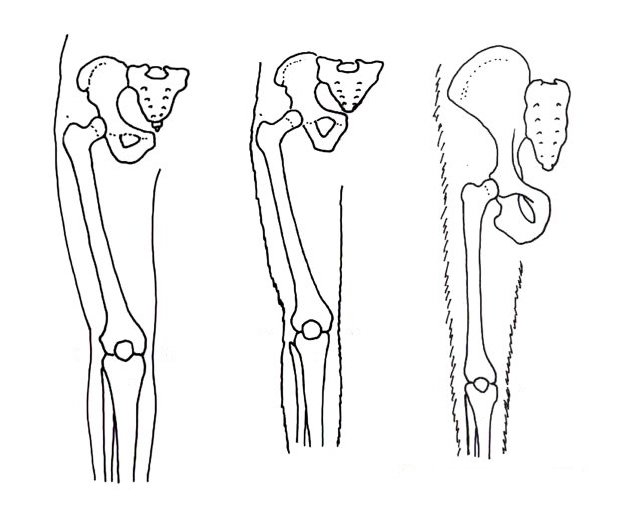
The oldest known hominin that verifiably possesses this trait is Orrorin tugenensis, an early hominin which lived in Kenya 6 million years ago. Orrorin is known from little fossil material, but what is known of it reveals lots about its locomotion and evolution. The most complete femur from this species is known as BAR 1002’00. This specimen is a left femur, and possesses an obvious bycondylar angle. Along with this, BAR 1002’00 has a thick epiphysis, meaning that Orrorin had the capability of bipedal locomotion, though it may not have been habitual.
Interestingly, the thickness of the epiphysis and the bycondylar angle are to a greater extent than what is found in later Australopithecus. This suggests that Orrorin was an off-branch species that evolved more bipedal traits before the rest of the hominins.

Right beneath the femur, the knee can also give a clue about how an organism walked. Human knee joints can fully extend and lock, allowing for the leg to extend straighter out. This trait is less significant, and it is unknown when it first appeared, but it could have come about anywhere from 4-2 million years ago.
At the very bottom of the body, the feet have several traits that allow for bipedal locomotion. Firstly, is the hallux bone. The hallux (big toe) in most primates is divergent, forming a more thumb-like shape. This makes the foot resemble a hand, allowing for the animal to be able to grasp branches with its hands and feet. Some hominins, such as Ardipithecus, possessed this trait, suggesting they were still spending time in the trees 4.4 million years ago, though the foramen magnum position suggests this genus still walked bipedally.

The divergent hallux isn’t lost until 4 million years ago with the genus Australopithecus, when hominins began using terrestrial bipedalism. This shows that it was a very quick change.
Aside from the hallux, the arches in the foot are also important for bipedal locomotion. Humans have three arches in our feet, the medial and lateral longitudinal arches, and the anterior transverse arch, while other apes have none. The transverse arch makes the foot stiffer, allowing it to bear more weight. This is useful for propulsion, allowing humans to walk bipedally more efficiently. The transverse arch specifically is significant as it contributes over 40% of the foot’s stiffness.
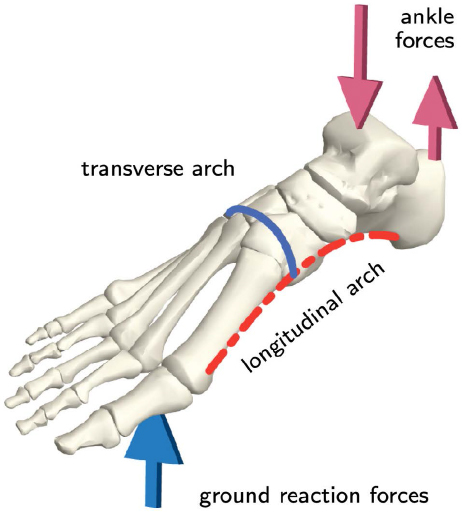
Hominin fossil feet are very rare, but there are several fossil specimens of hominin feet showing these arches. The first group to fully possess this trait are the Australopiths. Selam, the juvenile Australopithecus specimen mentioned earlier, has a surprisingly well preserved foot. The calcaneus (heel bone) of Selam is very well preserved and is very human-like. The heel morphologies place the tibia (shin bone) orthogonally (vertically over) foot as in modern humans, but the growth rate of the foot resembles more of other great apes. Other fossils of adult individuals of Australopithecus afarensis show morphologies more similar to adult humans.
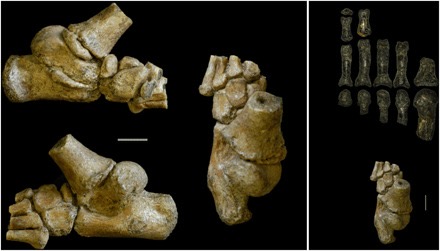
Another foot specimen, this time from Homo habilis, an early member of our genus from 2.5 million years ago shows similar, human-like morphology. The specimen, known as 0H 8, belonged to an adult individual, and is very similar to that of modern humans. The robustness of the foot and the foot arches are very similar to ours. These two finds show that from 4-2 million years ago, the foot morphology of hominins became much more adapted for terrestrial bipedalism and started resembling that of modern humans very quickly.
Now that we’ve looked at all the traits directly tied to bipedalism, let’s look at one which is indirectly related.
Because our tribe, Hominini, has specific traits, such as bipedal locomotion, if we find one fossil with other traits associated with hominins, it is reasonable to assume that they walked bipedally. Why would it have some traits but not the others? This isn’t the most reliable method, as evolution can often be very unpredictable, but it can be useful if it is all you have, and often times, it is all you have.
One of these traits is the size of the maxillary (upper) canines. In most apes, and primates in general, individuals have very large canines. These canines are often used for sexual and warning displays. Oftentimes for primates, the bigger the canines the better. The size of the canines also allows for a honing complex, where the upper canines are sharpened against the first premolars. This also resulted in the absence of a diastema, a gap in between the lower incisors and canines and upper canines that allowed for space for the large upper canine.
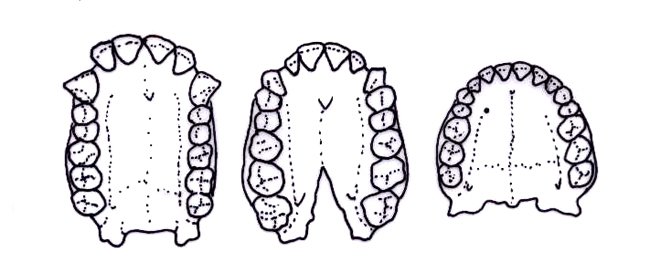
As hominins began evolving bipedal locomotion, their maxillary canines began being reduced. This allowed for non-honing chewing, giving our jaws more movement. Ardipithecus ramidus is the first species that shows significant canine reduction, and also has very little sexual dimorphism. It is around this time from 7-4 million years ago that hominins lost their large canines and began evolving bipedalism, so if a fossil ape maxilla is found with reduced canines, it very well could be a bipedal hominin, though you should also have precautions and never jump to conclusions.

There are other ways to tel how a species walked without physical fossil remains. Those are trace fossils, or footprints.
There are several preserved trackways from hominins showing their foot morphology and how they walked. The most famous of these trackways seems to be the Laetoli footprints from Tanzania.
Found in Tanzania, in a site known as Site G, roughly 70 hominin footprints are known, spanning about 88 feet. These tracks date to about 3.6 million years ago, and were likely made by 2, possibly 3 individuals of Australopithecus afarensis who were walking bipedally along a wet volcanic ashfall.
These tracks show a foot morphology very similar to modern humans, and a heel-strike method of walking, in which the heel touches the ground first, just like n modern humans, though 1.5 million year old tracks in Koobi Fora, Kenya, produced by Homo ergaster show an even more modern-like foot morphology and weight transfer, making the Laetoli footprints more primitive.
Research suggests that the hominins which produced the Laetoli footprints had stride lengths similar to modern humans, but there is debate on whether they used a bent-hip, bent-knee gait, similar to a chimpanzee, or an extended-hindlimb gait, similar to a modern human. This debate still seems to be unresolved, but there is lots of research going into it.

Conclusion:
After examining all the morphological characteristics necessary for bipedal locomotion, it is very easy to conclude how a hominin walked only from fossil remains. There are many traits, from the head to toes, many of which are insignificant or unexpected, but important nevertheless. Even without physical fossils, footprints produced by many different hominin species also can give a good idea on how these animals walked. It is important to give a good and honest look, as no matter what you have, it could very well give insight on the lifestyle, behavior, and evolution of our ancient ancestors.
Sources:
- Hagey, Travis. “Evolutionary Biomechanics”. Oxford Bibliographies, 01-12-23. https://www.oxfordbibliographies.com/display/document/obo-9780199941728/obo-9780199941728-0117.xml
- Plessner, Stephanie. “How Fish Swim”. Florida Museum. 03-27-18. https://www.floridamuseum.ufl.edu/discover-fish/fish/how-fish-swim/
- Gebo, D. L. (2013) Primate Locomotion. Nature Education Knowledge 4(8):1 https://www.nature.com/scitable/knowledge/library/primate-locomotion-105284696/
- Su, D. F. (2013) . Nature Education Knowledge 4(4):11 https://www.nature.com/scitable/knowledge/library/the-earliest-hominins-sahelanthropus-orrorin-and-ardipithecus-67648286/
- Blaxland, Beth, Dorey, Fran. “Walking on two legs-bipedalism”. Australian Museum. 02-10-20. https://australian.museum/learn/science/human-evolution/walking-on-two-legs-bipedalism/
- Williams, A. S., Zeng, I., Paton, J. G., Yelverton, C., Dunham, C., Ostrofsky, R. K., Shukman, S., Avilez, V. M., Eyre, J., Loewen, T., Prang, C. T., Meyer, R. M. (2022). Inferring lumbar lordosis in Neandertals and other hominins. PNAS Nexus, 1(1), pga005. https://doi.org/10.1093/pnasnexus/pgab005
- Wood, Matt. “3.3 million-year-old fossil reveals origins of the human spine”. UChicagoMedicine, 06-22-17. https://www.uchicagomedicine.org/forefront/biological-sciences-articles/3-3-million-year-old-fossil-reveals-origins-of-the-human-spine#:~:text=%22Selam%20has%20provided%20us%20the,had%20to%20take%20a%20trip
- Filiz, B. M., Toraman, F. N., Kutluk, G. M., Filiz, S., Doğan, K. S., Çakir, T., Yaman, A. (2021). Effects of lumbar lordosis increment on gait deteriorations in ambulant boys with Duchenne Muscular Dystrophy: A cross-sectional study. Brazilian Journal of Physical Therapy, 25(6), 749-755. 10.1016/j.bjpt.2021.05.001
- Gruss, T. L., Schmitt, D. (2015). The evolution of the human pelvis: changing adaptations to bipedalism, obstetrics and thermoregulation. Philosophical Transactions B, 370(1663), 20140063. 10.1098/rstb.2014.0063
- Friscia, Ton. “Evolutionary Anatomy – Walking Upright and Childbirth”. The Haps Blog, October 26, 2015. https://hapsblog.org/2015/10/26/evolutionary-anatomy-walking-upright-and-childbirth/
- Almécija, S., Tallman, M., Alba, M. D., Pina, M., Solà-Moyà, S., Jungers L. W. (2013). The femur of Orrorin tugenensis exhibits morphometric affinities with both Miocene apes and later hominins. Nature Communications, 4, 2888. https://doi.org/10.1038/ncomms3888
- Pickford, M., Senut, B., Gommery, D., Treil, J. (2002). Bipedalism in Orrorin tugenensis revealed by its femora. Comptes Rendus Palevol, 1(4), 191-203. https://doi.org/10.1016/S1631-0683(02)00028-3
- “Study Reveals an Early Tree-Dwelling Bipedal Human Ancestor was Similar to Ancient Apes and “Lucy but not Living Apes”. Stony Brook University, December 4, 2013. https://news.stonybrook.edu/newsroom/press-release/general/131204earlytreedwelling/
- Harcourt-Smith, E. H. W. (2010). The First Hominins and the Origins of Bipedalism. Evolution: Education and Outreach, 3, 333-340. https://doi.org/10.1007/s12052-010-0257-6
- Prang, C. T. (2019). The African ape-like foot of Ardipithecus ramidus and its implications for the origins of bipedalism. eLife, 8, e44433. https://doi.org/10.7554/eLife.44433
- Babu, D., Bordoni, B. (2022). Anatomy, Bony Pelvis, and Lower Limb: Medial Longitudinal Arch of the Foot. StatPearls (Internet). https://www.ncbi.nlm.nih.gov/books/NBK562289/
- Asghar, A.l, Naaz, S. (2021). The Role of Transverse Arch in Foot Stiffness and Its Clinical Implications. Anatomy and Biological Anthropology, 34(3), 103-104. https://doi.org/10.11637/aba.2021.34.3.103
- DeSilva, M. J., Gill., M. C., Prang, C. T., Bredella, A. M., Alemseged, Z. (2018). A nearly complete foot from Dikika, Ethiopia and its implications for the ontogeny and function of Australopithecus afarensis. 4(7), eaar7723. 10.1126/sciadv.aar7723
- Dorey, Fran. “Homo habilis”. The Australian Museum, 02-10-20. https://australian.museum/learn/science/human-evolution/homo-habilis/
- DeSilva, M. J., Zipfel, B., Arsdale, V. P. A., Tocheri, W. M. (2010). The Olduvai Hominid 8 foot: Adult or subadult? Journal of Human Evolution, 58(5), 418-423. https://doi.org/10.1016/j.jhevol.2010.03.004
- Delezene, K. L. (2015). Modularity of the anthropoid dentition: Implications for the evolution of the hominin canine honing complex. Journal of Human Evolution, 86, 1-12. https://doi.org/10.1016/j.jhevol.2015.07.001
- Suwa, G., Sasaki, T., Semaw, S., White, D. T. (2021). Canine sexual dimorphism in Ardipithecus ramidus was nearly human-like. PNAS, 118(49), e2116630118. https://doi.org/10.1073/pnas.2116630118
- Sanders, Robert. “New Ethiopian fossils are from 6-million-year-old hominid living just after split from chimpanzees”. UCBerkleyNews, 03-04-04. https://newsarchive.berkeley.edu/news/media/releases/2004/03/04_Akadab.shtml
- Dorey, Fran, Baxland, Beth. “Shorter jaws with smaller teeth”. The Australian Museum, 10-11-18. https://australian.museum/learn/science/human-evolution/shorter-jaws-with-smaller-teeth/
- “Laetoli Footprint Trails”. The Smithsonian Institution’s Human Origins Program” (ND). https://humanorigins.si.edu/evidence/behavior/footprints/laetoli-footprint-trails
- Raichlen, A. D., Gordon, D. A., Harcourt-Smith, H. E. W., Foster, D. A., Haas Jr., R. W. (2010). Laetoli Footprints Preserve Earliest Direct Evidence of Human-Like Bipedal Biomechanics. PLoS ONE, https://doi.org/10.1371/journal.pone.0009769
- “Footprints from Koobi Fora, Kenya”. The Smithsonian Institution’s Human Origins Program” (ND). https://humanorigins.si.edu/evidence/behavior/footprints/footprints-koobi-fora-kenya
- Raichlen, A. D., Pontzer, H., Sockol, D. M. (2008). The Laetoli footprints and early hominin locomotor kinematics. Journal of Human Evolution, 54(1), 112-117. https://doi.org/10.1016/j.jhevol.2007.07.005
- Reader, John. “Laetoli Footprints”. Science News, 01-01-1976. https://www.sciencenews.org/century/milestone/laetoli-footprints

I think I found an error above, near the beginning of the piece about locomotive methods.
“In organisms that walk bipedally, their foramen magnum is more posteriorly positioned, meaning it’s towards the back of the skull.”
I think you meant “quadrupedally” above where you have entered “bipedally”.
Johnny
LikeLike
Good catch, I don’t know how I missed that. I’ll try to fix it.
LikeLike