Introduction
The natural world is very often seen as dark and violent, a ‘dog-eat-dog world’. Though this isn’t always an accurate representation of nature, it isn’t entirely false either. Predatory animals will hunt and eat whatever they need to survive, to the extent of what they can hunt. We humans often like to think that we’re safe from these dangers, that we’re above the other animals and aren’t in danger from other predators we share this planet with. However, even a deep look into this will show that this isn’t the case, and never has been.
Shark Attacks are rare today in humans, but not completely unseen. Roughly 10 people a year are killed in shark attacks worldwide. Oftentimes these are surfers, who the sharks mistake as seals or sea lions, an animal they do commonly hunt. Archaeological evidence of shark attacks are even rarer. However, one example was described in 2021.
Evidence of a 3,000 year old shark attack has been uncovered from Japan, going back to the hunter-gatherer Jōmon period of the Japanese archipelago. This individual’s remains, known as Tsukumo 24, were buried at the Tsukumo site near Japan’s Seto Inland Sea, where modern shark attacks are relatively common to this day. From their remains, 790 perimortem injuries were found, characteristic of shark attacks. These injuries include deep bone gouges, punctures, cuts, and blunt force fractures. Most of the damage was on their pelvis, legs, shoulder, and arms. Along with this, their left hand and right leg were missing.
The right leg was present, but was upside down in the grave, and was not in articulation with the rest of the body. The distribution of the wounds suggest that the individual was alive during the attack, rather than being scavenged. The attack was fatal, and this individual was likely eventually killed due to blood loss and shock. The shark species responsible for the attack was most likely either a white shark (Carcharodon carcharias) or a tiger shark (Galeocerdo cuvier). Whatever remained of this individual’s body seems to have been retrieved and buried after.

This is a good example of humans being hunted in recent archaeological history, but even today, humans aren’t safe from other predatory animals. 600-800,000 human deaths are caused by tigers in Asia every year.
In Some places in India, leopards are massive threats, killing more people than all other cats combined. African cats aren’t as much of a threat, with cheetahs almost never hunting people, and lions rarely (but not never) hunting people, but it is a big problem in Asia.
A big reason for why predation is decreased in modern humans is because of our shelter and technology, protecting us from these dangers. Our other primate cousins however, do not have these luxuries, and face many more dangers. Not just our modern cousins however, but also our extinct ancestors and relatives. Examples of predation upon our ancestors are very prevalent throughout the fossil record.
The Predators of Extant and Extinct Primates
One of the most common predators of primates today are leopards. In Ethiopia, 7 encounters were recorded over a span of 6 years between geladas (Theropithecus gelada) and leopards (Panthera pardus).
Actual predation was rare, only being observed 1 time, but interactions occurred multiple times, typically ending with fear responses from the geladas, including distress calls and fleeing. Predation is rare towards baboon species due to their large social groups, known as multilevel societies, in which there are many individuals present. This suggests that having large social groups is advantageous to keep away predators. This lines up with the tendencies of leopards to hunt galadas in smaller social groups. Leopard hunting has also been observed in olive baboons (Papio anubis) as well, along with vervet monkeys (Chlorocebus pygerythrus).
Leopards hunt baboons most commonly at night. During the late Miocene to the Pliocene, around 3-7 million years ago, this would have provided a reason for our hominin ancestors to stay in the trees at night, to avoid these nocturnal predators. This didn’t save every hominin however.
SK 54
The skull cap fragment of a juvenile Paranthropus robustus, an extinct hominin relative, was uncovered in 1949, in a South African cave called Swartkrans, dating to 1.8-1.3 mya. On the back of the skull cap, near the lambdoid suture of the occipital bone, there are two puncture marks, matching the bite marks of a leopard. It seems that this individual was dragged away by the head by a leopard.
In the same deposit as SK 54, the mandible of a leopard was uncovered, known as SK 349. The canines of SK 349 fit in perfectly to the punctures of SK 54.The species of leopard that did this was a modern leopard (Panthera pardus), the same species that hunts modern primates today. This species arose in Africa during the Pleistocene, and would very soon migrate out of Africa to colonize places throughout Eurasia, with only the African variants still around today.

Off the ground, another predator of primates today and extinct hominins is the African crowned eagle.
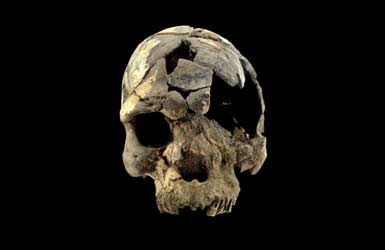
The Taung Child
A very big threat to African primates are predatory birds, specifically the African crowned eagle (Stephanoaetus coronatus). These eagles are very powerful and capable of killing mammals larger than themselves, including baboons.
In a 37 month study from Ngogo, Kibale National Park, 81% of the kill samples were monkeys. Redtail monkeys (Cercopithecus ascanius) were especially common, making up 66% of the identifiable monkey remains. Other research from the Tai Forest has collected 1,200 remains of animals hunted by these eagles. 669 of these were primate remains.
These eagles leave consistent taphonomic signatures, making them easily identifiable. The hind limbs and cranial bones often preserve well, while other parts, such as the ribs, vertebrae, carpals, and tarsals do not. Understanding the taphonomy of modern primates caused by crowned eagles, we can identify when this was the cause of death for extinct primates.
The skull of a juvenile Australopithecus africanus from South Africa, dating to 2-3.2 million years ago, who was about 3 years old when it died, unfortunately seems to have been killed by this species of eagle.
This skull is very significant for understanding human evolution, especially the brain development of our early ancestors, but is also important for understanding how these species interacted with their environments, and the threats they faced there.
The skull, known as the Taung Child, possesses talon marks in its orbits, identical to those left by crowned eagles in modern primates. Scratch marks are also present on the rest of the skull, including the frontal, temporal, parietal, and occipital bones. It seems that the Taung Child was carried off by a predatory eagle, and was eaten in its nest, where the skull would fall to the ground and be preserved.
OH 8
Another very common predator in Africa are crocodiles.
Crocodiles do hunt primates, but not as common as other large mammals. When they do hunt primates, they typically ambush them from the waters edge, just like what they do with other mammals. One example was observed in Indonesia in 1984.In this case, a juvenile crab-eating macaque (Macaca fascicularis) was ambushed by a crocodile when sitting at the edge of a river bank. This seems to be what happened to a juvenile Homo habilis.
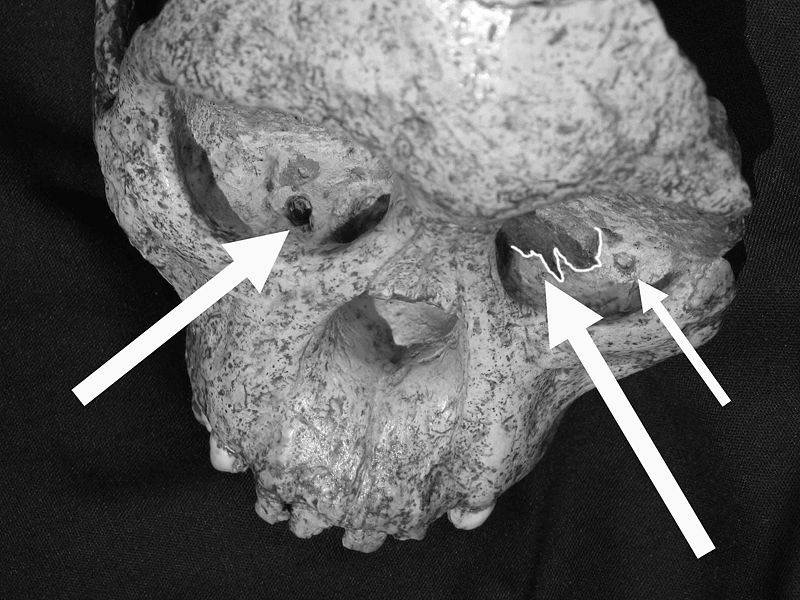
The OH 8 specimen is a foot belonging to a subadult or juvenile Homo habilis, based on the fusion of the metatarsals. This specimen is very important for understanding the locomotion of Homo habilis, as the arches in the feet are very significant for bipedal walking.The specimen is also possibly associated with the OH 7 mandible and the OH 35 leg bone. More interesting however, is the evidence of a crocodile attack in these fossils.
The foot, along with the possibly associated leg bone, bear extensive tooth marks indicative of a crocodile attack. The foot seems to have been disarticulated and torn from the rest of the body by the crocodile. The leg bone was found in a different deposit, and therefore may not belong to the same individual.
The fossils come from the FLK 22 FLK NN 1 fluvial sediments from the Olduvai paleo lake bed from Tanzania, dating to 1.8 mya. The crocodile that hunted OH 8 could have been a modern species of crocodile, but also could have been one of the many extinct crocodile species from the area. During the Pliocene and Pleistocene, there was a great diversity of crocodilians. One example of this is a species of horned crocodile from Oldovai Gorge, Tanzania, right where OH 8 was discovered. This species is characterized by large triangular ‘horns’ over its ears, along with a deep snout.
This species is very similar to modern crocodiles in the genus Crocodylus. The fossils also date to 1.8 million years ago, right alongside OH 8.This is the largest species of crocodile in the area, and very likely was the species that hunted our ancestors who lived alongside it.
Along with extinct crocodiles, there were many other extinct predators that are no longer alive today that would have been big threats to extinct hominins.

All the predators we’ve discussed so far, aside from the crocodile, are animals that are still alive today. However, there were plenty of animal species that our ancestors lived alongside that would have posed a major threat to them.
Ancient Predators
Most of the extinct predators from eastern and southern Africa were carnivorans, mainly big cats, but also including hyenas, canids (dogs), genets, mongooses, and extinct predatory otters. One site in Kenya preserved remains of all these animals, including orolutra sp., Enhydriodon (2 species), Genetta sp., Helogale sp., Homotherium sp., Dinofelis petteri, Felis sp., and Parahyaena howelli, dating to aout 5.3 mya. Some of these species are still around today, and not all would have hunted our ancestors, but the carnivoran diversity of eastern Africa during the Pliocene is nonetheless clear.

Down in South Africa, in Cooper’s Cave, a cave site known for Paranthropus robustus remains, many carnivoran remains are known as well.
These include mostly felids, such as Megantereon, Dinofelis, Panthera, Acinonyx, along with the small genera Caracal and Felis. Some of the most dangerous species from this site include the saber-toothed cats, Megantereon, Dinofelis, and Homotherium.
The saber-toothed cats belong to the group machairodontinae, a very diverse and successful felid lineage. Basal (early) members of this group include Promegantereon, Machairodus, Nimravides, Dinofelis, Metailurus, while more derived members include Megantereon, Amphimachairodus, Homotherium, Xenosmilus, and most famously, Smilodon.
The name eumachairodontinae has been proposed for these later species. Sub groups within this group include the tribes smilodontini, homotherini, and metalurini. The famous saber-teeth are exclusive to the later eumachairodonts, but other traits define the group machairodontinae as a whole. Multiple later machairodonts, or eumachairodonts, were in Africa at the same time as our earlier ancestors, and very likely hunted them.
Megantereon
Megantereon was a species of machairodontid that lived throughout Afro Eurasia, and even as far as North America, dating from about 2-4.2 million years ago.This species possesses at least 3 species (M. cultridens, M. falconeri and M. whitei) but possibly contains up to 13 species.There has been debate on whether different specimens represent different species, or are just examples of sexual dimorphism.
Their size averaged at about 3 feet long, though it varies slightly between species.Megantereon possessed large upper canines, but not as large as other related species, like Smilodon, giving it the name the “dirk-toothed cat”.This genus was very large and robust, suggesting that it was an ambush predator rather than a pursuit predator like a cheetah.
The species from Africa, Megantereon whitei, would have been the species that hunted our ancestors. Remains of this species are known mostly from Africa, though some fossils have been found as far as central Italy, showing evidence of faunal dispersal during the Plio-Pleistocene transition. Remains of African Megantereon, from northern Kenya dating to 3.5 mya, have been given the name Megantereon ekidoit, representing a possible second species of this genus in Africa.

Dinofelis
Another machairodont in Africa that lived alongside our ancestors was Dinofelis. Dinofelis is known from the same geographical range as Megantereon, dating from 5-1.2 mya. Belonged to the tribe metalurini, along with the other genera Metailurus, Adelphailurus, Stenailurus and Fortunictis.
There are up to 8 species of this genus, including D. cristata, D. diastemata, D. barlowi, D. paleoonca, D. darti, D. piveteaui, D. petteri, and D. aronoki, many of which come from Africa. The African species had a range from north Africa, all the way through down into South Africa.
Caves in South Africa possess Dinofelis remains alongside other hominins like Australopithecus africanus and Paranthropus robustus. Dinofelis had teeth very similar to a modern cheetah, specialized for slicing through flesh. Very likely hunted the hominins it lived alongside.

Homotherium
The last major big cat that hunted our ancestors is the infamous scimitar cat, Homotherium. Homotherium lived for the longest time out of the 3 big cats, living from 5 million years to 10,000 years ago, one of the most successful machairodonts. Along with a large temporal range, Homotherium had a great geographical range, living in the same places as Megantarion and Dinofelis. Later members of the species outside of Africa would have hunted large ice age mammals, like wooly mammoths, and even early Homo sapiens.
Homotherium was very large and robust, with powerful limbs and large teeth, and got up to 7 feet long, weighing 500 pounds.
Homotherium was a very diverse genus as well, with up to 15 species, such as Homotherium latidens, Homotherium ischyrus, Homotherium venezuelensis, and the African species, H. aethiopicum. Homotherium latidens is the species that would have interacted, and hunted modern humans throughout Eurasia.
Throughout the lifespan of this genus, it would have lived alongside our earliest ancestors, such as Australopithecus in Africa, all the way to modern humans in Eurasia, along with some of our other relatives, such as Homo heidelbergensis.

Relatives of cats, hyenas, would have also posed a threat to our ancestors, especially the hunting hyena, Chasmoporthetes.
Chasmaporthetes and Other Hyenas
Chasmaporthetes was a diverse genus of extinct hyena, which lived from 4.9 million years to 780,000 years ago, living throughout Afro Eurasia and into North America as well. The most well known species was Chasmaporthetes ossifragus, which lived in North America. It was a very fast and powerful hunter, preying upon other mammals like llamas, camels, deer, peccaries, and other small mammals. The species Chasmaporthetes lunensis ruled in Afro Eurasia. Chasmaporthetes often hunted human ancestors as well, including Homo erectus in Asia.
Hyenas are very prominent carnivores in Africa, but are mainly scavengers. It is possible that hyenas scavenged the hominin remains in the caves in South Africa. These hyenas would have been brown hyenas (Parahyaena brunnea) however, a living species, not the extinct Chasmaporthetes.
Outside of Africa, humans coexisted with other hyena species, such as the 46-33,000 year old Manot cave in Israel. The hyenas here were spotted hyenas (Crocuta crocuta). Bone assemblages from this cave show that the hyenas were mostly feasting upon fallow deer and mountain gazelles.
The humans here were also hunting other gazelles, but of different group sizes. The humans may have been hunting in more open areas due to their use of projectile weapons, but this also may be an example of niche partitioning amongst. humans and hyenas. Neanderthals in France were living alongside cave hyenas, exploiting different animal resources, just like the modern humans in Israel. The interactions between humans and hyenas in Europe weren’t always peaceful however.
In Guattari Cave, Italy, 9 Neanderthal individuals were uncovered. These Neanderthals weren’t living in this cave however, rather they were brought into it by hyenas. Many remains of Hyenas and Neanderthals are known from this cave, dating from 90-50,000 years ago. These hyenas were scavenging the bones of mostly adult males, along with one adult female and one juvenile. Along with Neanderthals, these hyenas were also scavenging on cows, rhinos, deer, bears, elephants, and horses.

Drimolen cave, South Africa, which dated to 2-1.5 million years ago, possesses remains of Dinofelis, modern leopards, and Chamaporthetes, showing that these African predators lived nearby to one another. There would have been some competition between these predators, especially the big cats that hunted in the South African environments, which were hunting the hominins in the area, along with baboon species. Chasmoporthetes had a more varied diet however.
Felids and hyenas were the most common predators in South Africa, with hyenas most commonly scavenging the hominins, just like what they do today. This was similar in eastern Africa, with felids being the primary predators.They would be hunting species like Paranthropus boisei and Australopithecus afarensis.
It is very clear that our ancient ancestors faced many dangers during their time from other animals. This begs the question, how did this affect our evolution? What evolutionary pressures did our predators produce? There are several ways these questions could be answered.
How Our Predators Shaped Us
There are multiple aspects of human biology and behavior that are results of our ancient predators. One big thing is our social structures.
Just like how other primates live in large social groups to avoid predators, it seems our ancestors did the same. The behaviors of other hominins are best compared to modern baboons as they live in similar environments. Primates that live in more open environments tend to have larger social groups as a predator avoidance tactic. Hominins more closely related to modern humans, like Homo ergaster, also lived in very sizable groups, having big implications for modern human sociality.
Comparisons with modern primates, mainly chimpanzees and baboons, suggests that Homo ergaster lived in large groups with many males, who would defend the group from predators. Defending against predators could have also developed into cooperative hunting, something very important for modern human behavior.
One of the reasons humans (and other primates as well) are so social and form such tight social groups could have started out as responses to the predation we faced in the African savanna. This is a human behavior shaped by predation, but there are also physical changes as well.
Primates have incredible vision compared to other mammals, including humans, at the cost of other senses, such as smell. One reason for this may be the dangers we faced in our past, mainly, snakes, big threats to not just primates, but all animals.
Snake bites are difficult to discern from fossils, but it is very likely that our ancestors faced them. Many dangerous snakes live in Africa, such as cobras, black mambas, vipers, and pythons, all of which pose threats to animals in Africa today, and no doubt would have done the same for our ancestors.

Snakes and placental mammals have evolved side by side, and snakes likely were one of the first major predators of these animals. Primate groups that have been more exposed to venomous snakes have larger parts of the brain associated with fear.
Snakes and primates have very interesting relationships. Many species of primates have been observed approaching, mobbing, killing, and even eating snakes. 26 primate species, including species which have attacked snakes, have been observed being killed and eaten by snakes. There is often aggression from both sides.
Similarly, studies on the Agta Negritos from the Philippines, have shown that these peoples have hunted and eaten pythons, along with deer, pigs, and wild monkeys. These same peoples have also been hunted by the same pythons. 26% of adult males have been reported to have survived python attacks, and 6 deaths were reported between 1934-1973. Though not super common, predation from snakes on primates is prevalent enough to have a possible evolutionary significance.
Humans and other primates are very sensitive to images of snakes, more so than any other animal. Having increased vision would have been very beneficial for detecting the camouflaged snakes. Mosaic snakes, equivalent to camouflage snakes, stand out to people much more than any other animal. Snakes have even appeared as more threatening than guns and knives, things that are much more dangerous in the present day.
This suggests that humans have an instinctual fear response to snakes, as a result of visual adaptations to avoid snakes. This idea is known as snake detection theory. This theory is further backed up by the fact that other primates have similar responses to snake images.
Vervet and macaque monkeys respond very strongly to partially exposed images of snakes and images of snake scales more than any other animal, mainly lizards and birds. A natural fear response towards snakes is not only present in humans, but many primates, showing that it’s an ancestral instinct, seemingly from the earliest primates which evolved alongside snakes.
The predation from these threatening reptiles shaped our brains and senses, having effects in our brains even to this day.
Conclusion
Though today, we are mostly safe from the predators that hunt the animals that live alongside us, this hasn’t always been the case. Evidence of predation on humans and our ancestors is found from as recent as 3,000 years ago to as long ago as 3 million years ago. From water bound sharks, to land ridden big cats, and to airborne eagles, our ancestors have always been in danger. These predators have had big effects on our evolution as well, altering our sociality, and even possibly our vision. As sad as it is to think about these ancient apes being hunted, we should also be grateful for it, as without these past dangers, we wouldn’t be the way we are today.
Sources
- Tobin, Barry. “Sharks”. Australian Institute of Marine Science (ND). https://www.aims.gov.au/docs/projectnet/sharks-02.html
- Hensley, Justin. “Big cats and you”. LITFL, 06-03-20. https://litfl.com/big-cats-and-you/#:~
- White, A. J., Burgess, H. J., Nakatsukasa, M., Hudson, J. M., Pouncett, J., Kusaka, S., Yoneda, M., Yamada, Y., Schulting, J. R. (2021). 3000-year-old shark attack victim from Tsukumo shell-mound, Okayama, Japan. Journal of Archaeological Sciences: Reports, 38, 103065. https://doi.org/10.1016/j.jasrep.2021.103065
- Weisberger, Mindy. “Oldest-known shark attack discovered in 3,000-year-old skeleton with 800 injuries”. Live Science, 06-25-21. https://www.livescience.com/shark-attack-victim-3000-years-ago.html
- Baxland, B., Dorey, F. “Do we know how some early human ancestors died?” Australian Museum, 03-12-21. https://australian.museum/learn/science/human-evolution/how-do-we-know-how-they-died/
- Pomeroy, R. “What Hunted Ancient ‘Humans’”. RealClear Science, 04-24-19. https://www.realclearscience.com/blog/2019/01/24/what_hunted_ancient_humans.html
- Lin, B., Foxfoot, R. I., Miller, M. C., Venkatamaran, V. V., Kerby, T. J., Bechtold, K. E., Kellogg, S. B., Nguyen, Fashing, J. P. (2020). Leopard predation on gelada monkeys at Guassa, Ethiopia. American Journal of Primatology, 82(2), e23089. https://doi.org/10.1002/ajp.23098
- Isbell, A. L., Bidner, R. L., Cleave, V. K E., Matsumoto-Oda, A., Crofoot, C. M. (2018). GPS-identified vulnerabilities of savannah-woodland primates to leopard predation and their implications for early hominins. Journal of Human Evolution, 118, 1-13. https://doi.org/10.1016/j.jhevol.2018.02.003
- Tawane, Mirriame. “Fossils and the evidence they provide.” Creative Feel, 04-08-21. https://creativefeel.co.za/2021/04/fossils-and-the-evidence-they-provide-the-tale-of-sk-54-and-sk-349/
- Paijmans, A. L. J., Barlow, A., Förster, W. D., Henneberger, K., Meyer, M., Nickel, B., Nagel, D., Havmøller, W. R., Baryshnikov, F. G., Joger, U., Rosendahl, W., Hofreiter, M. (2018). Historical biogeography of the leopard (Panthera pardus) and its extinct Eurasian populations. BMC Ecology and Evolution. 18, 156. https://doi.org/10.1186/s12862-018-1268-0
- Sanders, J. W., Trapani, J., Mitani, C. J. (2003). Taphonomic Aspects of Crowned Hawk-Eagle Predation on Monkeys. Journal of Human Evolution, 44(1), 87-105. https://doi.org/10.1016/S0047-2484(02)00196-3
- McGraw, S. W., Cooke, C., Shultz, S. (2006). Primate remains from African crowned eagle (Stephanoaetus coronatus) nests in Ivory Coast’s Tai Forest: Implications for primate predation and early hominid taphonomy in South Africa. American Journal of Biological Anthropology, 131(2), 151-165. https://doi.org/10.1002/ajpa.20420
- “Taung Child”. The Smithsonian Institution’s Human Origins Program, 08-30-22. https://humanorigins.si.edu/evidence/human-fossils/fossils/taung-child
- Berger, R. L., McGraw, S. W. (2007). Further evidence for eagle predation of, and feeding damage on, the Taung child. South African Journal of Science. 103, 11-12. http://www.scielo.org.za/scielo.php?pid=S0038-23532007000600013&script=sci_arttext
- Berger, R. L. (2006). Brief communication: Predatory bird damage to the Taung type-skull of Australopithecus africanus Dart 1925. American Journal of Biological Anthropology, 131(2), 166-168. https://doi.org/10.1002/ajpa.20415
- “Crowned Eagle”. eBird (ND). https://ebird.org/species/crheag1
- Galdikas, F. M. B., Yeager, P. C. (1984). Brief report: Crocodile predation on a crab-eating macaque in Borneo. American Journal of Primatology, 6(1), 49-51. https://doi.org/10.1002/ajp.1350060106
- DeSilva, M. J., Zipfel, B., Van Arsdale, P. A., Tocheri, W. M. (2010). The Olduvai Hominid 8 foot: Adult or subadult? Journal of Human Evolution, 58(5), 418-423. https://doi.org/10.1016/j.jhevol.2010.03.004
- Susman, L. R., Patel, A. B., Francis, J. M., Cardoso, V. F. H. (2011). Metatarsal fusion pattern and developmental morphology of the Olduvai Hominid 8 foot: Evidence of adolescence. Journal of Human Evolution, 60(1). 58-69. https://doi.org/10.1016/j.jhevol.2010.09.004
- Njau, K. J., Blumenschine, J. R. (2012). Crocodylian and mammalian carnivore feeding traces on hominid fossils from FLK 22 and FLK NN 3, Plio-Pleistocene, Olduvai Gorge, Tanzania. Journal of Human Evolution, 63(2), 408-417. https://doi.org/10.1016/j.jhevol.2011.05.008
- Blumenschine, J. R., Stanistreet, G. I., Njau, K. J., Bamford, K. M., Masao, T. F., Albert, M. R., Stollhofen, H., Andrews, P., Prassack, A. K., McHenry, J. L., Fernández-Jalvo, Y., Camili, L. E., Ebert, I. J. (2012). Environments and hominin activities across the FLK Peninsula during Zinjanthropus times (1.84 Ma), Olduvai Gorge, Tanzania. Journal of Human Evolution, 63(2), 364-383. https://doi.org/10.1016/j.jhevol.2011.10.001
- “OH 8”. The Smithsonian Institution’s Human Origins Program. 07-01-22. https://humanorigins.si.edu/evidence/human-fossils/fossils/oh-8
- Brochu, A. C., Njau, J., Blumenschine, J. R., Densmore, D. L. (2010). A New Horned Crocodile from the Plio-Pleistocene Hominid Sites at Olduvai Gorge, Tanzania. PLoS ONE, 5(2), e9333. https://doi.org/10.1371/journal.pone.0009333
- “Pliocene (Pt 10): Before There Were Zebras.” Synapsida, 04-17-16. https://synapsida.blogspot.com/2016/04/pliocene-pt-10-before-there-were-zebras.html#
- Werdelin, L., Lewis, E. M. (2020). A contextual review of the Carnivora of Kanapoi. Journal of Human Evolution, 140, 102334. https://doi.org/10.1016/j.jhevol.2017.05.001
- Pester, Patrick. “Lion-size otters prowled Ethiopia 3 million years ago”. Live Science, 09-13-22. https://www.livescience.com/lion-sized-otter-unearthed-ethiopia
- O’Regan, J. H., Steininger, C. (2017). Felidae from Cooper’s Cave, South Africa (Mammalia: Carnivora). Geodiversitas, 39(2), 315-332. https://doi.org/10.5252/g2017n2a8
- Christiansin, P. (2012). Phylogeny of the sabertoothed felids (Carnivora: Felidae: Machairodontinae). Cladistics, 29(5), 543-669. https://doi.org/10.1111/cla.12008
- “Megantereon”. Prehistoric Wildlife (ND). http://www.prehistoric-wildlife.com/species/m/megantereon.html
- Palmqvist, P., Torregrosa, V., Pérez-Claros, A. J., Martínez-Navarro, B., Turner, A. (2006). A re-evaluation of the diversity of Megantereon (Mammalia, Carnivora, Machairodontinae) and the problem of species identification in extinct carnivores. Journal of Vertebrate Paleontology, 27(1), 160-175. https://doi.org/10.1671/0272
- Christiansen, P., Adolfssen, S. J. (2007). Osteology and ecology of Megantereon cultridens SE311 (Mammalia; Felidae; Machairodontinae), a sabrecat from the Late Pliocene – Early Pleistocene of Senéze, France. Zoological Journal of the Linnean Society, 151(4), 833-884. https://doi.org/10.1111/j.1096-3642.2007.00333.x
- Sardella, R., Petrucci, M., Rook, L. (2008). The African species Megantereon whitei from the Early Pleistocene of Monte Argentario (South Tuscany, Central Italy). Comptes Rendus Palevol, 7(8), 601-606. https://doi.org/10.1016/j.crpv.2008.09.009
- Palmqvist, P. (2015). On the presence of Megantereon whitei at the South Turkwel Hominid Site, northern Kenya. Journal of Paleontology, 76(5), 928-930. 10.1666/0022-3360(2002)0762.0.CO;2
- Madurell, Malapeira, J., Rodríguez-Hidalgo, A., Aouraghe, H., Haddoumi, H., Lucenti, B. S., Oujaa, A., Saladié, P., Bengamra, S., Marín, J., Souhir, M., Farkouch, M., Mhamdi, H., Aissa, M. A., Werdelin, L., Chacón, G. M., Sala-Ramos, R. (2021). First small-sized Dinofelis: Evidence from the Plio-Pleistocene of North Africa. Quaternary Science Reviews, 265, 107028. https://doi.org/10.1016/j.quascirev.2021.107028
- Piegl, Linda, Bothma, Bianca. “Dinofelis – hominid hunter or misunderstood feline?”. Maropeng, 12-20-11. https://www.maropeng.co.za/news/entry/dinofelis_hominid_hunter_or_misunderstood_feline
- Strauss, Bob. “Homotherium”. ThoughtCo, 11-04-19. https://www.thoughtco.com/homotherium-same-beast-1093219
- Antón, M., Salesa, J. M., Galobart, A., Tseng, J. Z. (2014). The Plio-Pleistocene scimitar-toothed felid genus Homotherium Fabrini, 1890 (Machairodontinae, Homotherini): diversity, palaeogeography and taxonomic implications. Quaternary Science Reviews, 96, 259-168. https://doi.org/10.1016/j.quascirev.2013.11.022
- Antón, M., Galobart, A., Turner, A. (2005). Co-existence of scimitar-toothed cats, lions and hominins in the European Pleistocene. Implications of the post-cranial anatomy of Homotherium latidens (Owen) for comparative palaeoecology. Quaternary Science Reviews, 24(10-11). 1287-1301. https://doi.org/10.1016/j.quascirev.2004.09.008
- “The American Hyena (Chasmaporthetes ossifragus)”. Georgia Before People (ND). https://markgelbart.wordpress.com/2013/10/01/the-american-hyena-chasmaporthetes-ossifragus/
- Arriaza, C. M., Aramendi, J., Maté-González, A. M., Yravedra, J., Stratford, D. (2021). The hunted or the scavenged? Australopith accumulation by brown hyenas at Sterkfontein (South Africa). Quaternary Science Reviews, 273, 107252. https://doi.org/10.1016/j.quascirev.2021.107252
- Orbach, M., Yeshurun, R. (2021). The hunters or the hunters: Human and hyena prey choice divergence in the Late Pleistocene Levant. Journal of Human Evolution, 160, 102572. https://doi.org/10.1016/j.jhevol.2019.01.005
- Dusseldorp, L. G. (2013). Neanderthals and Cave Hyenas: Co-existence, Competition or Conflict? Zooarchaeology and Modern Human Origins, 191-208, https://doi.org/10.1007/978-94-007-6766-9_12
- Luntz, Stephen. “Remains Of Nine Neanderthals Eaten By Hyenas Found In Italian Cave”. Live Science, 05-10-21. https://www.iflscience.com/remains-of-nine-neanderthals-eaten-by-hyenas-found-in-italian-cave-59651
- O’Regan, J. H., Menter, C. G. (2009). Carnivora from the Plio-Pleistocene hominin site of Drimolen, Gauteng, South Africa. Geobios, 42(3), 329-350. https://doi.org/10.1016/j.geobios.2009.03.001
- Lee-Thorp, J., Thackeray, F. J., van der Merwe, N. (2000). The hunters and the hunted revisited. Journal of Human Evolution, 3(6), 565-576. https://doi.org/10.1006/jhev.2000.0436
- Fourvel, J., Thackeray, Brink, S. J., O’Reagan, H., Braga, J. (2018). Taphonomic interpretations of a new Plio-Pleistocene hominin-bearing assemblage at Kromdraai (Gauteng, South Africa). Quaternary Science Reviews, 190, 81-97. https://doi.org/10.1016/j.quascirev.2018.04.018
- Aramendi, J., Arriaza, C. M., Yravedra, J., Maté-González, A. M., Ortega, C. M., Courtenay, A. L., González-Aguilera, D., Gidna, A., Mabulla, A., Baquedano, E., Domínguez-Rodrigo, M. (2019). Who ate OH80 (Olduvai Gorge, Tanzania)? A geometric-morphometric analysis of surface bone modifications of a Paranthropus boisei skeleton. Quaternary International, 517, 118-130. https://doi.org/10.1016/j.quaint.2019.05.029
- Badenhorst, S. (2018). Possible predator avoidance behaviour of hominins in South Africa. South African Journal of Science, 114, 7-8. http://dx.doi.org/10.17159/sajs.2018/a0274
- Willems, P. E., van Schaik, P. C. (2017). The social organization of Homo ergaster: Inferences from anti-predator responses in extant primates. Journal of Human Evolution, 109, 11-21. https://doi.org/10.1016/j.jhevol.2017.05.003
- “The Largest 10 Snakes in Africa”. A-Z Animals (ND). https://a-z-animals.com/blog/the-10-largest-snakes-in-africa/
- Isbell, A. L. (2006). Snakes as agents of evolutionary change in primate brains. Journal of Human Evolution, 51(1), 1-35. https://doi.org/10.1016/j.jhevol.2005.12.012
- Headland, N. T., Greene, W. H. (2011). Hunter–gatherers and other primates as prey, predators, and competitors of snakes. PNAS, 108(52), E1470-E1474. https://doi.org/10.1073/pnas.1115116108
- Kawai, N., He. H. (2016). Breaking Snake Camouflage: Humans Detect Snakes More Accurately than Other Animals under Less Discernible Visual Conditions. PLoS ONE, 11(10), e0164342. https://doi.org/10.1371/journal.pone.0164342
- Van Strien, W. J., Isbell, A. L. (2017). Snake scales, partial exposure, and the Snake Detection Theory: A human event-related potentials study, Scientific Reports, 7, 46331. https://doi.org/10.1038/srep46331