
Introduction
During the Pleistocene epoch, the world was full of human species, all throughout Africa, Europe, and Asia, and eventually, North America. Human species were interacting with, living with, and even breeding with each other. Some had vast continent-wide homes, while some migrated far away into new unexplored lands, such as obscure islands in Indonesia or the Philippines. It was a fascinating time, and is one heavily studied by anthropologists today. There is lots of fossil and genetic evidence providing clear pictures of most of these species, but there are also many gaps in our knowledge.
The many fossils we have are very useful for understanding this time in human history, but also can cause lots of confusion, mainly in terms of taxonomy. In a previous article (https://worldofpaleoanthropology.org/2023/08/23/understanding-the-muddle-in-the-middle-hominins-from-the-pleistocene-guest-post-by-mekhi/), I discussed the Pleistocene hominins from Europe and some from Africa, and the confusion that comes with them, but recent research has added more confusion to the other species, this time living farther east, in Asia.
There were many hominin species in Asia, and they arguably cause more confusion than what is seen in Africa and Europe. To understand this, we first need to go through all the known human species from the area.
Hominin Diversity and Distribution During the Pleistocene of Asia
Animals often have large geographical ranges, and live in more than one place. Leopards (Panthera pardus) for example, have a huge range, mostly living in Africa, but also living in Europe and Asia, having the biggest range of any big cat today. Some species of humans had similarly large ranges. Obviously, Homo sapiens today have a massive range, living pretty much across the whole globe, but even other extinct species had large ranges as well. It is important to note a big difference between humans and leopards in this comparison though, which is that though leopards have a broad geographic range, the species is split into different subspecies in different areas, whereas other extinct human species are typically not.
A good example of broad geographic ranges is Homo heidelbergensis from Africa and Europe, though there has been contention on whether the African populations were the same species or not. Another species, one more relevant to this topic, is Homo neanderthalensis, or the Neanderthals, known from both Europe and (sparsely) from Asia.
Neanderthals in Asia
Neanderthals were mostly settled in Europe, but were also present in Asia, mainly western Asia and the middle east, but sites containing their remains are also known farther east from Russia. Because it can sometimes be tricky to identify whether a specimen actually belonged to a Neanderthal or not, especially one so far from home, genetic analysis is often used. Genetic research on hominin remains in Russia shows that Neanderthals were present as far as the Altai region of Siberia.
A cave called Chagyrskaya in this region is very famous for Neanderthals. This site, dated to ~60,000 years ago (kya), is not only a great example of the Neanderthal remains in Asia, with many tools, Neanderthal bones, and bones of other animals are known from it, but it is also a prime spot for Neanderthal genetics.
Genetic research has shown that this population was relatively isolated, and had less than 60 individuals present, which is still large for Neanderthal standards. The genetic material from this cave is also exceptionally well preserved. The genome of one individual was sequenced for example, showing that she was female and closely related to the other known Neanderthals from western Asia.
Another cave, very close to Chagyrskaya, also documents Neanderthals in Russia; it is known as Okladnikov cave. Nearby to both these caves, housing a very unknown species only found in Asia, is Denisova cave, known for the Denisovans.

Denisovans
The Denisovans were a species of late genus Homo, very similar to Neanderthals, that lived in east Asia ~500-40 kya. Though our knowledge about this species is still growing, they’re still a cause of great confusion, mostly due to the fact that we have very, very few fossils. We have no skeletons, no skulls, nothing, just a few teeth and bone fragments. Almost everything we know about them is known just from genetic material. What we actually can learn about Denisovan morphology comes mostly from the dental remains.
The morphology seen in Denisovan teeth is mostly unique from that of Neanderthals and Homo sapiens, showing their designation as a separate species. The teeth do show similarities with Asian Homo erectus, but the overall morphology is mostly unique.
Though the Denisovans were most likely a distinct species, no true species name has been given due to the lack of fossil material. Species names like Homo altaiensis or Homo denisova may be used in the future.
The Asian Neanderthals and Denisovans also inhabited the same region, at around the same time. Neanderthals and Denisovans were sister species, and shared a recent common ancestor that was itself a sister species to Homo sapiens. Similar to how people can have some levels of Neanderthal DNA within their genomes, people today can also have some Denisovan DNA, with present-day Melenesians having up to 4-6% Denisovan DNAin their genomes. Much of the genetic material we do have of Denisovans to compare this comes from the few physical remains we actually have from them.
Denisovan genetic material is known from multiple remains. The most important and famous example of this may be the individual known as “Denny”. This individual, known from the fragmentary ‘Denisova 11’ remains, has very well preserved genetic material. From these remains, we know the sex of this individual, which is female, her eye color, which is brown, and most importantly, who her parents were. Her father was a Denisovan, but her mother was a Neanderthal, making her a hybrid. She also was ~13 years of age.

This shows that not only were Neanderthals living with and interacting with Denisovans, but they were also breeding with them. This suggests that Neanderthals were in this region by 120 kya, even as old as potentially 190 kya. Very little is known about Denisovan biology, but more is known about their culture.
Evidence of fire use, mainly charcoal and ashes in the sediments, has been found in Denisova cave, suggesting these people were using fire, which isn’t very surprising. Stone tools have also been uncovered in the cave, giving insight to their technology. Multiple stages of occupation can be found from these tools, each from the different and distinct sedimentary layers containing different tool assemblages from different times. Layers 11.3, 13, 14, and 15 come from the middle paleolithic, and layers 11.1 and 11.2 from the upper paleolithic.
Also from the upper paleolithic layers, specifically the initial upper paleolithic, are lots of bone tools and decorative ornaments. These bone tools consist of an eyed needle and awe from the bones of large mammals. The bone ornaments, or jewelry, include beads, pendants, plaques, rings, a button/fastener, and decorated bones and bracelets.
These bone tools show that the Denisovans were smart, and could take advantage of multiple resources. It also shows that they were culturally advanced, and could make jewelry, though simple, to decorate their bodies. It has long been thought jewelry was something unique to modern Homo sapiens, but we now have evidence that other human species including both Denisovans and Neanderthals were doing it too.

The Denisovans were and still are an odd and confusing species, but more species were causing more confusion down in southeast Asia at this time. One such species was Homo floresiensis, also known as the ‘hobbit’ species.
Homo floresiensis
Homo floresiensis at first sounds like a normal hominin species, living on the island of Flores in Indonesia from ~190-50 kya. This species was one of the Pleistocene hominins in Asia, along with Neanderthals, Denisovans, and all the other species that will be discussed later, but there is something unique, and very confusing about this species.
Though very human-like, clearly in the genus Homo, this species was very primitive, especially in the fact that the adults were only ~1 meter (~3 feet) tall. This short stature of this strange species has caused great confusion, and a few different hypotheses have been proposed to explain it.
At first, the fossils, found in Liang Bua cave, were interpreted as pathological modern humans, humans who suffered from microcephaly (a decrease in the size of the head). However, this hypothesis was very quickly shown to be false. The skull of H. floresiensis and that of a microcephalic human are very different, and H. floresiensis lacks the expected features for a person with this condition. This gave a need for new hypotheses. H. floresiensis clearly wasn’t a modern human, nor was it ancestral to modern humans; it was a strange ‘hobbit’ species living on a secluded island in Indonesia.
This idea of an off-branching species was very important at the time; it was commonly thought that human evolution was a straight line of progression, with the only branching members of the lineage being groups like the robust australopiths (Paranthropus) and Neanderthals. This species showed the true diversity of hominins that lived before modern humans that weren’t actually ancestral to modern humans.
The next hypothesis for the origins of these hobbits was that they were a subset of Homo erectus, a species that inhabited the nearby island of Java at the time, that simply suffered from insular dwarfism, a common evolutionary phenomenon that occurs when species are trapped on isolated islands with limited resources. This would explain why H. floresiensis was so short and why it was there, but it would not explain the rest of the morphology.
Even early on in the research of H. floresiensis, scientists noticed that the species had many basal, or primitive characteristics, traits shared with earlier African hominins such as Australopithecus and early Homo. Some of the obvious primitive traits include its short stature, but it also had a small brain volume (~380 cubic centimeters), and limb proportions similar to that of earlier hominins. However, it still had some derived features shared with later species like Homo erectus, such as a flatter, more orthognathic face, and more derived teeth.
Cladistic analysis suggests that H. floresiensis arose sometime around the time of Homo habilis and Homo rudolfensis, the two earliest known members of the genus Homo, though it is possible that it came sometime after early Homo. The origin of H. floresiensis is still highly debated; both ideas are very plausible and have support, but the main consensus right now seems to be that this species is a descendant of early hominin species from Africa, and just happened to end up nearby to where H. erectus would inhabit.
This however does bring up another question, the question of how this species even got to the island in the first place. Flores is an island, and has not recently been in contact with other land masses, unlike other Indonesian islands like Java. This means that H. floresiensis had to cross some waterways to get there. This isn’t actually too implausible. Much of the native wildlife of the island, from mammals to reptiles, seem to have come to the island from neighboring islands, such as Java, Sahul, and Sulawesi. Some of the larger animals, such as large reptiles and the extinct elephant relative Stegodon likely could have swam or even floated, while the smaller mammals, such as rodents, may have rafted there. These are all also species that H. floresiensis ate and butchered.
This rafting hypothesis easily applies to H. floresiensis. We can never know the true path the hobbits took to get there, but they very well may have rafted, likely unintentionally, to the island they would call home. They would have a long time of success on the island after their arrival, but went extinct at some point, and their extinction offers even further confusion.
Natural disasters, such as tsunamis and volcanic events are common there, and definitely had effects on them, but they don’t seem to have wiped them out. A large volcanic eruption is known to have occurred on Flores ~12 kya, but this was too late in time, as H. floresiensis seems to have been extinct by this point. Their disappearance seems to have occurred ~50 kya, which matches exactly with the known arrival date of Homo sapiens on the island.
For a brief period, our species seems to have coexisted with the hobbits, and unfortunately, we may have led to their extinction. The origins and disappearance of these hobbits adds more confusion to what already was a confusing time and place, but they weren’t the only confusing species in the area.

Homo luzonensis
While Homo floresiensis was living in Indonesia, another confusing species was living nearby in the Philippines, called Homo luzonensis. Similar to the Denisovans, H. luzonensis is known from very little fossil material, though we have more from this species than we have from the Denisovans. Unlike the Denisovans however, we have no genetic material from H. luzonensis. No full skeletons have been found either, unlike H. floresiensis. Only 13 remains dated to 67 kya have been uncovered. Overall, very little is known about this species.
The remains from this species include mostly teeth, some finger and toe bones, and the femur of a young child. All the remains come from the same stratigraphic layer of Callao Cave, in the island of Luzon. Though very small in numbers, the fossils we do have can tell us some things, or at least give us hints. The teeth may be the most useful of the remains for learning about this species.
Out of the 13 remains, 7 of them were teeth, 5 of which seem to have belonged to the same individual. These 5 teeth make up the holotype of the species, another reason why they have a species name and the Denisovans don’t. The teeth are primitive and small, and similar to those of H. floresiensis.
The small size of the teeth suggests that H. luzonensis may have had a small body size, as tooth size often reflects body size, though this isn’t always the case. The metatarsals and metacarpals in the feet and hands are also very small and primitive, resembling H. floresiensis and early members of the genus like Homo habilis. Despite the size of the teeth, they also share many traits with Homo erectus. This could suggest that H. luzonensis descended from H. erectus and potentially suffered some insular dwarfism, though it is possible that it was a descendant of earlier hominins, like what is thought for H. floresiensis.
Like H. floresiensis, the origins of H. luzonensis are unknown and heavily debated, though, also like H. floresiensis, whoever they descended from, they likely got to the island via rafting, as Luzon has never been recently in contact with any other landform. If not from earlier hominin species, these two island based human species must have descended from H. erectus, the only other hominin species living in the area.
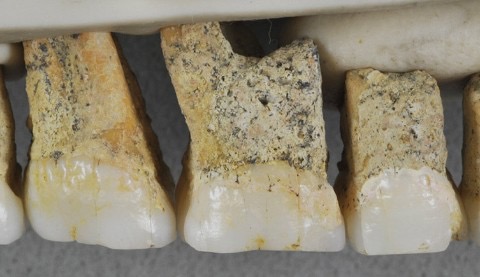
Homo erectus
Homo erectus was perhaps the most successful human species, at least for its time. The oldest H. erectus fossils are known from 1.8 million years ago (mya), to as recent as ~100 kya. H. erectus arose out of Africa, and migrated all throughout Eurasia, settling mostly in southeast Asia.
The site known as Sangiran, in the Solo basin on the island of Java Indonesia has revealed over 40 H. erectus specimens, making it perhaps the most important locality for H. erectus in Indonesia. Sangiran is one of the 3 main sites known for Homo erectus remains in Eurasia, with the others being Dmanisi in eastern Europe and Zhoukoudian in China. The earliest of these fossils in Indonesia are about 1.5m years old, which is very close to the 1.8m years old remains in Dmanisi, the oldest H. erectus remains out of Africa. This shows that they spread and migrated very quickly.
This population in Indonesia however lived for a very long time after that; they’re seemingly the last surviving group of H. erectus. 12 H. erectus skull caps, along with 2 leg bones, uncovered from Ngandong, central Java, near the Solo River, represent the last known H. erectus population to have lived, along with the most derived population. These fossils lived between 117-108 kya, showing truly just how long H. erectus lived as a species.
This brings up the question of how did H. erectus reach Java in the first place? The answer to that is glaciation. During the Pleistocene, as old as 2.6 mya, glaciations resulted in drastic drops in sea level, as much as 72-120 meters (~236-393 feet) below current sea level in some cases. This resulted in many land bridges across Indonesia, allowing for faunal migration to the islands. This led to the colonization of Java by many different animals by about 2 mya, including mammoths and likely H. erectus, though it is likely that they still had to cross some water bodies to get there.
When they arrived at the island, they were introduced to an environment of moist grasslands and open woodlands, with riverine landscapes and large streams, along with long annual dry seasons. Aside from Indonesia, H. erectus is also commonly known from China, mainly at a site called Zhoukoudian.
Zhoukoudian represents ~80 individuals, the most number of H. erectus individuals known from anywhere, along with ~100,000 stone tools. This population arrived there long after others arrived in Indonesia, living there from ~750-200 mya, though their occupation there seems to have been sporadic.
Unfortunately, by 108k years, the all-mighty Homo erectus had gone extinct. It is hypothesized that their extinction was caused by climate and environmental changes in the region. The vast and open environments they migrated into, found in both Indonesia and China, transitioned to a wet and crowded rainforest, which may have ended their reign. However, we likely will never know what truly wiped them out, as it is hard to point at the exact causes of small extinctions.
H. erectus is the cause of little confusion, being one of the better understood species from the Pleistocene. However, there was another human population living in China that is incredibly unknown, unique and confusing, more so than any of the other species of the time, making the name the ‘muddle in the middle’ especially well deserved.

Homo longi
The next species in Asia is called Homo longi. This species is the most recently named human species, being first described in 2021. Even though it was first described in 2021, the skull, known as the Harbin skull, was originally discovered in 1933 during a construction project in the Hebei Province of northern China, although the exact locality is unknown. The skull was dated to ~146 kya.
The skull itself is one of the most well preserved hominin crania from the Pleistocene of Asia; it is only missing most of its teeth and possesses damage to its left zygomatic arch. It is also very large. It has a large cranial capacity of ~1,420 cubic centimeters. Some of its morphology is similar to that of Homo sapiens, such as its large cranial capacity, short face, and small cheekbones, but it also possesses some other unique morphology. This includes a low cranial vault, pronounced brow ridges, large molars, and alveolar prognathism below the nose.
It also shares features with Homo heidelbergensis and Neanderthals. This is a unique and strange combination of traits. This set of traits is also seen in other hominin crania from China, that seem to represent a unique and unknown human population or species living in China during this time, that includes Homo longi. As it turns out, there is a lot of hominin fossil material from the Pleistocene from all throughout China, and these fossils are the most confusing part of human evolution in the Pleistocene of Asia.

The Mysterious Hominins of Pleistocene China
Much of the hominin fossil record of Pleistocene China, just as it is as a whole, is teeth. Below is a chart showing different locations in China where hominin teeth were uncovered, spanning a temporal range of 990-15 kya.
| Location of Discovery | Dating | Number of Teeth |
| Meipu, southern China | 990-780 KYA | 4 |
| Yiyuan, eastern China | 770-126 KYA | 7 |
| Longtan, Cave, Hexian, eastern China | 412 KYA | 10 |
| Panxian Dadong, southern China | 300-130 KYA | 4 |
| Tongzi, southern China | 172-240 KYA | 4 |
| Luna Cave, southern China | 126-70 KYA | 2 |
| Dushan Cave, southern China | 15 KYA | 1 |
Only a few of the dental remains have been assigned a species, such as the teeth from Luna cave, which clearly belong to Homo sapiens. The morphology of these teeth is also interesting. Many of the teeth have Homo erectus-like features, while other traits appear more modern. This mosaic morphology in the dentition is similar to the mosaic morphology of the rest of the bodies, especially the skulls.
This mosaic nature, the mix of archaic and modern features has caused some to suggest that many of these remains belong to hybrid individuals, hybrids of archaic and modern human species, such as H. erectus and H. sapiens. This idea has especially been proposed for some of the postcranial material that has been uncovered, mainly femora.
The skulls from this time and place, as mentioned, also display strange morphology, and have been under great research lately, revealing a lot about the hominins living in China. A collection of well preserved hominin crania, including the Harbin skull of Homo longi, make up the best set of hominin remains from this time, and are the most useful and important. Well preserved hominin crania are known from 5 sites, Harbin, Dali, Jinnisuan, Maba, and maybe most importantly, Hualongdon.
Dali
The Dali skull, from the Shaanxi Province of northwest China, was originally uncovered in 1978. This skull has been estimated to be ~200k years old. The specimen is mostly complete, only having some distortion and a few missing pieces, including a big chunk of the parietal bone.
The face of the Dali skull resembles that of Homo sapiens, but aside from the face, the skull resembles other African and eastern Eurasian Pleistocene hominins. Overall, it most greatly resembles the skulls of early H. sapiens of north Africa and the Levant region. This combination of traits from multiple hominins is a consistent trend throughout all these skulls.
The Dali skull may show that during the Pleistocene, many different human species and populations in Asia were breeding with one another, creating a ‘braided stream’, with gene flow from different groups creating unique morphology in different populations. At its extremes, this idea is known as the ‘multiregional hypothesis’, and is in contention with the theory of ‘out of Africa’.


Jinniushan
The next important specimen is not not just a skull, but a whole partial skeleton and cranium from northeast China. This is very rare in the hominin fossil record from this time, as most specimens are either just a skull, or fragmentary remains like teeth or small postcranial (body) material. This is not the case for the Jinniushan specimen, which consists of a skull, along with an arm bone (left ulna), a pelvic bone (left innominate), 6 vertebrae, ribs, and bones from the hands and the feet.
The skull itself is fairly complete, but was broken during its excavation in 1984. What is missing from the skull is mostly from the frontal, parietal, and occipital regions. The dating of this specimen has been somewhat controversial, but current estimates place it at roughly ~200 kya, similar in time to Dali. Other fossils come from the same strata as the Jinniushan specimen, including bears, wolves, deer, and rhinos.
Morphologically, the skull shares some similarities with Dali, along with a mix of Homo erectus and Homo sapiens features. They both possess a small mastoid process, similar parietal bones, and a shared location of the maximum cranial breadth. There are many differences too, including smaller brow ridges, a shallower supraorbital sulcus, etc. The cranial capacity of the Jinnushan skull is roughly 1,400 cubic centimeters, which is very large, and around the same size as modern H. sapiens.
The presence of well preserved postcranial material provides a rare case in which the encephalization quotient (the size of the brain compared to the body) can be seen. In Jinniushan, the encephalization quotient is 4.15, which is very close to the predicted encephalization quotient for other pleistocene hominins from this area. As for the rest of the body, this individual’s body mass was estimated to be 78.6 kg (~172 lbs), and the body height to be 168 cm (~5 ½ ft). It also had shorter limbs than expected.
All these body proportions are consistent with adaptations for colder weather, in which species adapt to be larger and have shorter limbs to conserve body warmth. This is known as Allen’s rule, and is also seen in other hominin species such as Neanderthals. The climate in China during the Pleistocene is not extensively studied, but research has revealed a climate transition, known as the Mid-Pleistocene climate transition (MPT), characterized by increased monsoon and glaciation cycles from ~1.2m to 700 kya. This occurred before the Jinniushan specimen was around, but the presence of these features may suggest similar conditions during its time.
The foot of the Jinniushan specimen is also interesting. It possessed some derived features, such as an increased stability of the medial longitudinal arch, but it also had several primitive features, such as lower arches and a less stable hallucal metatarsophalangeal joint. These traits provide a glimpse into the stride of this pleistocene hominin; they suggest that this individual had a slightly different stride than what is seen in modern H. sapiens.
Other details about the Jinniushan specimen include that it was likely female based off of the pelvis, and was ~16-20 years of age, making her relatively young. Stone tools and traces of fire have also been reported alongside the Jinniushan specimen.
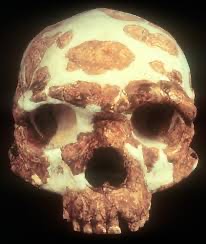
Maba
After Jinniushan in this skull set comes the Maba skull. This skull, from ~300-130 kya is very fragmentary, consisting of a skull cap, a right orbit, and a portion of the nasal bone. Along with this, it possesses pronounced brow ridges and a heavily curved frontal bone, more so than Chinese Homo erectus. The brow ridges are also similar in shape and size to European Neanderthals.
The thickness of the cranial vault is also similar to that of modern Asian Homo sapiens. Just as with all the other crania discussed so far, Maba possesses a strange mosaic of features shared between multiple other hominin species. It is also somewhat unique from Dali and Jinniushan, having a more prognathic face. The cranial capacity of Maba is about 1,300 cubic centimeters, within the ranges of Homo heidelbergensis, Neanderthals, and H. sapiens. There are a few other hominin remains aside from the cranium as well, consisting of 5 teeth and a partial mandible, though they seem to not be associated with the Maba cranium, as they dated to ~237 kya, and some of the teeth are associated with H. sapiens instead.


The last strange skull that will be discussed is one of the more recently described specimens, like Harbin. It may be one of the more important specimens known from this time and place; it is the HLD6 Hualongdong skull.
Hualongdong
The taxonomic placement of all these skulls is very confusing. They’re all unique in their own ways, and all have mosaic characteristics, combining multiple traits from several other hominins. One commonly thought idea is that these skulls, or at least for some of them, are Denisovans. They’re in the right place and time, but this could never be confirmed.
We have no confirmed Denisovan cranial material; there is nothing to definitively back up the claim that these Chinese skulls represent Denisovans, and no genetic material from these specimens to compare. There is the Denisovan partial mandible, Xiahe, but no cranium, and all of the skulls discussed here are crania, but have no associated mandibles, at least up until Hualongdong.
The specimen known as HLD 6, from Hualongdong eastern China, is a ~300k year old juvenile hominin skull and mandible, along with some dental remains. The skull’s face resembles that of other Asian hominin crania, including Dali, Jinniushan, Maba, and even Homo erectus. The face of the skull is very flat, like Homo sapiens, though this could also be because the individual was young, only ~13-15 years of age; face shape changes lots in hominin ontogeny. Overall, the cranium fits in well with the other mosaic skulls from this time and place. However, the mandible is the more important part of the specimen.

The mandible is broadly characterized by a robust mandibular corpus (the body of the mandible), and a more gracile mandibular symphysis and ramus (the front and back ends of the mandible). It also lacks a chin, showing that it can’t belong to H. sapiens. It does however share many other traits with other pleistocene hominin mandibles, including a pronounced alveolar planum, a superior transverse torus, a pronounced endocondyloid crest, and a well-developed medial pterygoid tubercle.
It shares lots in common with other Pleistocene hominins from throughout Eurasia. This is significant as it is one of the only complete hominin mandibles from China, only going alongside Penghu 1 from Taiwan (~190-10 kya), Xujiayao 14 from northern China (~120 kya), and the Denisovan Xiahe mandible (~160 kya).
Despite also being from China, the HLD6 mandible differs greatly from these other mandibles. This is significant as no other of the Chinese crania discussed so far also have associated mandibles. Because the HLD6 cranium is so similar to the other crania, it is reasonable to assume that its mandible is similar as well. If the mandibles are similar, it means that all the mandibles are distinct from the other Chinese mandibles.
This is especially important for the Xiahe mandible. If the Xiahe mandible is distinct from the other mandibles, then the cranium may be too. This could imply that the Denisovan cranium and these other Chinese crania are distinct, showing that these hominins may not be Denisovans. This especially applies to Homo longi, as it has been long speculated to be Denisovan. This could be a huge discovery, but it is very important to note that this claim is entirely based on logical speculation, and will need significant further, physical evidence to be proven.

What Are These Skulls?
So now we know what the skulls (probably) aren’t, what exactly are they? These fossils are clearly not modern Homo sapiens, and also shouldn’t be grouped in as Homo erectus either, despite its presence in the region at this time. One species that these hominins have sometimes been grouped under is Asian Homo heidelbergensis.
Homo heidelbergensis already has had a history of confusion when it comes to many specimens being grouped into it that may not necessarily belong there. Though many of the African and European hominin specimens fit into H. heidelbergensis, the Asian specimens seem to be overall distinct.
The 2 main reasons why these Asian specimens are sometimes grouped into H. heidelbergensis are 1) some apomorphic (derived) traits seen in Asia are also found in European H. heidelbergensis, and 2) Homo sapiens didn’t descend from Asian Homo erectus, so it must be H. heidelbergensis instead.
Some of the shared traits include very heavily increased encephalization, and the presence of canine fossa, which is found in several of the Chinese specimens and also the fossils from Gran Dolina Spain, associated with Homo antecessor. This shows that the Chinese specimens may represent a group closer to H. heidelbergensis cladistically, and farther from H. erectus.
However, despite these shared traits, there are many distinguishing traits as well. These traits include the orientation of the frontosphenoidal process of the zygomatic bone, the upper facial height, shovel shaped incisors, the presence of interparietal bones, and agenesis of the 3rd molars. Some H. heidelbergensis do possess some of these traits, such as interparietal bones, but they’re uncommon.
Overall, it is unlikely that the Chinese specimens represent H. heidelbergensis, but they may be related to it. Another common idea for the classification of these specimens is placing them in a new species. Homo longi is the only one of the Chinese crania that has an actual assigned species, but other species names have been proposed for other specimens, such as Homo daliensis and Homo mabaensis. This however, is counterproductive; the Pleistocene is already very confusing in terms of hominin taxonomy with lots of very confusing taxa; adding more taxa will only muddle it up more.
It might not be terrible to group them all under one diverse species, such as M. mabaensis, but all this would do would be changing what we call the fossils, and wouldn’t do much for our actual understanding. It has been proposed that the safest course of action in terms of what to call these specimens is simply to refer to them as archaic Homo sapiens until further analysis clears up the muddle in Asia.
Conclusion
This mysterious group of hominins clearly were a distinct group, likely closely related to H. heidelbergensis and other Pleistocene hominins. Classifying these hominins is difficult, as there isn’t any consistent set of traits that define them. The only truly defining trait is the increasing morphological diversity throughout all of them. It is also unknown how these hominins ended up where they did. There are 3 hypotheses for how they got to China, but nothing for certain.
The first idea is that they got there via the Chao Phraya river basin; the second is that they got there through east Vietnam, and the 3rd and last idea is that they traveled along the coast of southern Asia and Myanmar. These are all plausible ideas, but none of them have enough evidence to back them up to make them certain.
Until more evidence and research comes along, the Pleistocene of Asia will remain confusing, and, just like with Europe, further research and discoveries, such as new fossils and genetic evidence are our best hope to resolve this confusing time.
Sources
- “Where Do Leopards Live? In the Wild, Of Course – Right?” africafreak (ND). https://africafreak.com/where-do-leopards-live
- Dorey, Fran. “Homo heidelbergensis”. The Australian Museum, 06-28-21.https://australian.museum/learn/science/human-evolution/homo-heidelbergensis/
- Dorey, Fran. “Homo neanderthalensis”. The Australian Museum, 06-28-21. https://australian.museum/learn/science/human-evolution/homo-neanderthalensis/
- Krause, J., Orlando, L., Serre, D., Viola, B., Prüfer, K., Richards, P. M., Hublin, J., Hänni, C., Derevianko, P. A., Pääbo, S. (2007). Neanderthals in central Asia and Siberia. Nature, 499, 902-904. https://doi.org/10.1038/nature06193
- Salazar-García, C. D., Power, C. R., Rudaya, N., Kolobova, K., Markin, S., Krivoshapkin, A., Henry, G. A., Richards, P. M., Viola, B. (2021). Dietary evidence from Central Asian Neanderthals: A combined isotope and plant microremains approach at Chagyrskaya Cave (Altai, Russia). Journal of Human Evolution, 156, 102589. https://doi.org/10.1016/j.jhevol.2021.102985
- Mafessoni, F., Grote, S., de Filippo, C., Pääbo, S. (2020). A high-coverage Neandertal genome from Chagyrskaya Cave. PNAS, 11(26): 15132-15136. https://doi.org/10.1073/pnas.2004944117
- Dobrovolskaya, V. M., Tuinov, V. A. (2013). The Neanderthals of Okladnikov Cave Altai: Environment and Diet Based on Isotopic Analysis. Archaeology, Ethnology, and Anthropology of Eurasia. 41(1): 78-88. https://doi.org/10.1016/j.aeae.2013.07.007
- Dorey, Fran. “The Denisovans”. The Australian Museum, 20-04-20. https://australian.museum/learn/science/human-evolution/the-denisovans/
- Zubova A. V., Chikisheva T .A., Shunkov M. V. (2017). The Morphology of Permanent Molars from the Paleolithic Layers of Denisova Cave. Archaeology, Ethnology & Anthropology of Eurasia. 45(1): 121-134. https://doi.org/10.17746/1563-0110.2017.45.1.121-134
- Jacobs Z., Li, B., Shunkov, V. M., Kozlikin, B. M., Bolikovskaya, S. N., Agadjanian, K. A., Uliyanov, A. V., Vasiliev, K. S., O’Gorman, K., Derevianko, P. A., Roberts, G. R. (2019). Timing of archaic hominin occupation of Denisova Cave in southern Siberia. Nature, 565, 594-599. https://doi.org/10.1038/s41586-018-0843-2
- Reich, D., Green, E. R., Kircher, M., Krause, J., Patterson, N., Durand, Y. E., Viola, B., Briggs, W. A., Stenzel, U., Johnson, F. L. P., Maricic, T., Good, M. J., Marques-Bonet, T., Alkan, C., Fu, Q., Mallick, S., Li, H., Meyer, M., Eichler, E. E., Stoneking, M., Richards, M., Talamo, S., Shunkov, V. M., Derevianko, P. A., Pääbo, S. (2010). Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature, 468, 1053-1060. https://doi.org/10.1038/nature09710
- Slon, V., Mafessoni, F., Vernot, B., de Filippo, C., Grote, S., Viola, B., Hajdinjak, M., Peyrégne, S., Nagel, S., Brown, S., Douka, K., Higham, T., Kozlikin, B. M., Shunkov, V. M., Derevianko, P. A., Kelso, J., Meyer, M., Prüfer, K., Pääbo, S. (2018). The genome of the offspring of a Neanderthal mother and a Denisovan father. Nature, 561, 113-116. https://doi.org/10.1038/s41586-018-0455-x
- Morely, W. M., Goldberg, P., Uliyanov, A. V., Kozlikin, B. M., Shunkov, V. M., Derevianko, P. A., Jacobs, Z., Roberts, G. R. (2019). Hominin and animal activities in the microstratigraphic record from Denisova Cave (Altai Mountains, Russia). Scientific Reports, 9, 13785. https://doi.org/10.1038/s41598-019-49930-3
- Shunkov, V. M., Kozlikin, B. M., Derevianko, P. A. (2020). Dynamics of the Altai Paleolithic industries in the archaeological record of Denisova Cave. Quaternary International, 559, 34-46. https://doi.org/10.1016/j.quaint.2020.02.017
- Shunkov, V. M., Fedorchenko, U. A., Kozlikin, B. M., Derevianko, P. A. (2020). Initial Upper Palaeolithic ornaments and formal bone tools from the East Chamber of Denisova Cave in the Russian Altai. Quaternary International, 559, 47-67. https://doi.org/10.1016/j.quaint.2020.07.027
- O’Donoghue, Saskia. “Ancient human DNA found in 25,000-year-old tooth jewelry”. Euronews, 08-05-2023. https://www.euronews.com/culture/2023/05/08/ancient-human-dna-found-in-25000-year-old-tooth-jewellery
- Dorey, Fran. “Homo floresiensis”. The Australian Museum, 23-12-19. https://australian.museum/learn/science/human-evolution/homo-floresiensis/
- Groves, C. (2007). The Homo floresiensis Controversy. Hayati Journal of Biosciences, 14(4): 123-126. https://doi.org/10.4308/hjb.14.4.123
- Argue, D., Morwood, J. M., Sutkina, T., Jatmiko, Saptomo, W. E. (2009). Homo floresiensis: a cladistic analysis. Journal of Human Evolution, 57(5): 623-639. https://doi.org/10.1016/j.jhevol.2009.05.002
- Aiello, C. L. (2010). Five years of Homo floresiensis. American Journal of Biological Anthropology, 142(2): 167-179. https://doi.org/10.1002/ajpa.21255
- Brumm, A., Aziz, F., van den Bergh, D. G., Morwood, J. M., Moore, W. M., Kurniawan, I., Hobbs, R. D., Fullagar, R. (2006). Early stone technology on Flores and its implications for Homo floresiensis. Nature, 441, 624-628. https://doi.org/10.1038/nature04618
- Dennell, W. R., Louys, J., O’Reagan, J. H., Wilkinson, M. D. (2014). The origins and persistence of Homo floresiensis on Flores: biogeographical and ecological perspectives. Quaternary Science Reviews, 96, 98-107. https://doi.org/10.1016/j.quascirev.2013.06.031
- Davis, Josh. “Homo luzonensis: new species of ancient human discovered in the Philippines”. Natural History Museum, 04-11-19. https://www.nhm.ac.uk/discover/news/2019/april/new-species-of-ancient-human-discovered-in-the-philippines.html
- Détroit, F., Mijares, S. A., Corny, J., Daver, G., Zanolli, C., Dizon, E., Robles, E., Grün, R., Piper, J. P. (2019). A new species of Homo from the Late Pleistocene of the Philippines. Nature, 568, 181-186. https://doi.org/10.1038/s41586-019-1067-9
- Zanolli, C., Kaifu, Y., Pan, L., Xing, S., Mijares, S. A., Kullmer, O., Schrenk, F., Corny, J., Dizon, E., Robles, E., Détroit, F. (2022). Further analyses of the structural organization of Homo luzonensis teeth: Evolutionary implications. Journal of Evolution, 163, 103124. https://doi.org/10.1016/j.jhevol.2021.103124
- Roberts, L. D., Jarić, I., Lycett, J. S., Flicker, D., Key, A. (2023). Homo floresiensis and Homo luzonensis are not temporally exceptional relative to Homo erectus. Journal of Quaternary Science, 38(4): 463-470. https://doi.org/10.1002/jqs.3498
- Dorey, Fran. “Homo erectus”. The Australian Museum, 16-10-20. https://australian.museum/learn/science/human-evolution/homo-erectus/
- Zaim, Y., Ciochon, L. R., Polanski, M. J, Grine, E. F., Bettis, III, A. E., Rizal, Y., Franciscus, G. R., Larick, R. R., Aswan, H. M., Eaves, L. K., Marsh, E. H. (2011). New 1.5 million-year-old Homo erectus maxilla from Sangiran (Central Java, Indonesia). Journal of Human Evolution, 61(4): 364-376. https://doi.org/10.1016/j.jhevol.2011.04.009
- Widianto, H., Noerwidi, S., Hascaryo, T. A. (2023). New Hominin calvaria discovery from Grenzbank Layer of Sangiran Dome (Java, Indonesia): The last archaic Homo erectus lived in Java. L’Anthropologie, 127(3): 103165. https://doi.org/10.1016/j.anthro.2023.103165
- Rizal, Y., Westaway, E. K., Zaim, Y., van den Bergh, D. G., Bettis, III, A. E., et al. (2019). Last appearance of Homo erectus at Ngandong, Java, 117,000–108,000 years ago. Nature, 577, 381-385. https://doi.org/10.1038/s41586-019-1863-2
- Widianto, H., Noerwidi, S. (2023). Long journey of Indonesian Homo erectus: Arrival and dispersal in Java Island. L’Anthropologie, 127(3): 103167. https://doi.org/10.1016/j.anthro.2023.103167
- Bettis, III, A. E., Milius, K. A., Carpenter, J. S., Larick, R., Zaim, Y., Rizal, Y., Ciochon, L. R., Tassier-Surine, A. S., Suminto, M. D., Bronto, S. (2009). Way out of Africa: Early Pleistocene paleoenvironments inhabited by Homo erectus in Sangiran, Java. Journal of Human Evolution, 56(1): 11-24. https://doi.org/10.1016/j.jhevol.2008.09.003
- Choi, Q. Charles. “The Final Days of Homo Erectus”. Science Insider.12-18-19. https://www.insidescience.org/news/final-days-homo-erectus#
- Ni, X., Ji, Q., Wu, W., Li, Q., Grün, R., Stringer, C. (2021). Massive cranium from Harbin in northeastern China establishes a new Middle Pleistocene human lineage. The Innovation, https://doi.org/10.1016/j.xinn.2021.100130
- Liu, W., Schepartz, A. L., Xing, S., Miller-Antonio, S., Wu, X., Trinkaus, E., Martinón-Torres, M. (2013). Late Middle Pleistocene hominin teeth from Panxian Dadong, South China. Journal of Human Evolution, 64(5): 337-355. https://doi.org/10.1016/j.jhevol.2012.10.012
- Xing, S., Martinón-Torres, M., Bermúdez de Castro, M. J., Zhang, Y., Fan, X., Zheng, L., Huang, W., Liu, W. (2014). Middle Pleistocene Hominin Teeth from Longtan Cave, Hexian, China. PLoS ONE, https://doi.org/10.1371/journal.pone.0114265
- Xing, S., Martinón-Torres, M., Deng, C., Shao, Q., Wang, Y., Luo, Y., Zhou, X., Pan, L., Ge, J., Bermúdez de Castro, M. J., Liu, W. (2021). Early Pleistocene hominin teeth from Meipu, southern China. Journal of Human Evolution, 151, 102924. https://doi.org/10.1016/j.jhevol.2020.102924
- Xing, S., Martinón-Torres, M., Bermúdez de Castro, M. J. (2019). Late Middle Pleistocene hominin teeth from Tongzi, southern China. Journal of Human Evolution, 130, 96-108. https://doi.org/10.1016/j.jhevol.2019.03.001
- Liao, W., Xing, S., Li, D., Martinón-Torres, M., Wu, X., Soligo, C., Bermúdez de Castro, M. J., Wang, W., Liu, W. (2019). Mosaic dental morphology in a terminal Pleistocene hominin from Dushan Cave in southern China. Science Reports, 9, 2347. https://doi.org/10.1038/s41598-019-38818-x
- Xing, S., Sun, C., Martinón-Torres, M., Bermúdez de Castro, M. J., Han, F., Zhang, Y., Liu, W. (2016). Hominin teeth from the Middle Pleistocene site of Yiyuan, Eastern China. Journal of Human Evolution, 95, 33-54. https://doi.org/10.1016/j.jhevol.2016.03.004
- Bae, J. C., Wang, W., Zhao, J., Huang, S., Tian, F., Shen, G. (2014). Modern human teeth from Late Pleistocene Luna Cave (Guangxi, China). Quaternary International, 354, 169-183. https://doi.org/10.1016/j.quaint.2014.06.051
- Pan, L., Dumoncel, J., Mazurier, A., Zanolli, C. (2020). Hominin diversity in East Asia during the Middle Pleistocene: A premolar endostructural perspective. Journal of Human Evolution, 148, 102888. https://doi.org/10.1016/j.jhevol.2020.102888
- Pan, L., Dumoncel, J., Mazurier, A., Zanoli, C. (2019). Structural analysis of premolar roots in Middle Pleistocene hominins from China. Journal of Human Evolution, 136, 102669. https://doi.org/10.1016/j.jhevol.2019.102669
- Curnoe, D., Ji, X., Liu, W., Bao, Z., Taçon, C. S. P., Ren, L. (2015). A Hominin Femur with Archaic Affinities from the Late Pleistocene of Southwest China. PLoS ONE, https://doi.org/10.1371/journal.pone.0143332
- Wei, P., Weng, Z., Carlson, J. K., Cao, B., Jin, L., Liu, W. (2021). Late Pleistocene partial femora from Maomaodong, southwestern China. Journal of Human Evolution, 155, 102977. https://doi.org/10.1016/j.jhevol.2021.102977
- “Dali”. PeterBrown-Anthropology (ND). https://www.peterbrown-palaeoanthropology.net/dali.html
- Athreya, S., Wu, X. (2017). A multivariate assessment of the Dali hominin cranium from China: Morphological affinities and implications for Pleistocene evolution in East Asia. American Journal of Biological Anthropology, 164(4): 679-701. https://doi.org/10.1002/ajpa.23305
- Wu, X., Athreya, S. (2012). A description of the geological context, discrete traits, and linear morphometrics of the Middle Pleistocene hominin from Dali, Shaanxi Province, China. American Journal of Biological Anthropology. 150(1): 141-157. https://doi.org/10.1002/ajpa.22188
- “Jinniushan”. PeterBrown-Anthropology (ND). https://www.peterbrown-palaeoanthropology.net/jinniush.html
- “Jinniushan Man Skull (replica)”. National Museum of China, 08-27-21. https://en.chnmuseum.cn/collections_577/collection_highlights_608/archaeological_discoveries_609/202109/t20210927_251612.html
- Rosenberg, R. K., Zuné, L, Ruff, B. C. (2006). Body size, body proportions, and encephalization in a Middle Pleistocene archaic human from northern China. PNAS, 103(10): 3552-3556. https://doi.org/10.1073/pnas.0508681103
- Yang, S., Yue, J., Zhou, X., Storozum, M., Huan, F., Deng, C., Petraglia, D. M. (2020). Hominin site distributions and behaviours across the Mid-Pleistocene climate transition in China. Quaternary Science Reviews, 248, 106614. https://doi.org/10.1016/j.quascirev.2020.106614
- Lu, Z., Meldrum, J. D., Huang, Y., He, J., Sarmiento, E. E. (2011). The Jinniushan hominin pedal skeleton from the late Middle Pleistocene of China. HOMO, 62(6): 389-401. https://doi.org/10.1016/j.jchb.2011.08.008
- “Maba”. PeterBrown-Anthropology (ND). https://www.peterbrown-palaeoanthropology.net/maba.html
- Wu, X., Bruner, E. (2016). The endocranial anatomy of maba 1. American Journal of Biological Anthropology. 160(4): 633-643. https://doi.org/10.1002/ajpa.22974
- Xiao, D., Bae, J. C., Shen, G., Delsin, E., Jin, H. H. J., Webb, M. N., Qiu, L. (2014). Metric and geometric morphometric analysis of new hominin fossils from Maba (Guangdong, China). Journal of Human Evolution, 74, 1-20. https://doi.org/10.1016/j.jhevol.2014.04.003
- Bower, Bruce. “‘Dragon Man’ skull may help oust Neanderthals as our closest relative”. Science News, 06-25-21. https://www.sciencenews.org/article/dragon-man-skull-homo-longi-human-evolution-neandertals
- Wu, X., Pei, S., Cai, Y., Liu, W. (2019). Archaic human remains from Hualongdong, China, and Middle Pleistocene human continuity and variation. PNAS, 116(20): 9820-9824. https://doi.org/10.1073/pnas.1902396116
- Wu, X., Pei, S., Cai, Y., Tong, H., Xing, S., Jashashvili, Carlson, J. K., Liu, W. (2021). Morphological description and evolutionary significance of 300 ka hominin facial bones from Hualongdong, China. Journal of Human Evolution, 161, 103052. https://doi.org/10.1016/j.jhevol.2021.103052
- Chang, C., Kaifu, Y., Takai, M., Kono, T. R., Grün, R., Matsu’ura, S., Kinsley, L., Lin, L. (2015). The first archaic Homo from Taiwan. NaturePorfolio, 27(6): 6037. 10.1038/ncomms7037
- Wu, X., Trinkaus, E. (2014). The Xujiayao 14 Mandibular Ramus and Pleistocene Homo Mandibular Variation. Comptes, Rendus, Palevol, 13(4): 333-341.
- https://doi.org/10.1016/j.crpv.2013.10.002
- Wu, X., S. Pei, Cai, Y., Tong, H., Zhang, Z., Yan, Y., Xing, S., Martinón-Torres, M., Bermúdez de Castro, M. J., Liu, W. (2023). Morphological and morphometric analyses of a late Middle Pleistocene hominin mandible from Hualongdong, China. Journal of Human Evolution, 182, 103411. https://doi.org/10.1016/j.jhevol.2023.103411
- Sáez, Roberto. “A new sister group of the sapiens lineage: on the skulls of Hualongdong and Harbin”. Nutcracker Man, 10-25-21. https://nutcrackerman.com/2021/10/25/nuevo-grupo-hermano-linaje-sapiens-craneos-hualongdong-harbin/
- Bae, J. C. (2010). The late Middle Pleistocene hominin fossil record of eastern Asia: Synthesis and review. American Journal of Biological Anthropology, 143(S51): 75-93. https://doi.org/10.1002/ajpa.21442
- Liu, W., Athreya, S., Xing, S., Wu, X. (2022). Hominin evolution and diversity: a comparison of earlier-Middle and later-Middle Pleistocene hominin fossil variation in China. Philosophical Transactions of the Royal Society B, 377(1848). https://doi.org/10.1098/rstb.2021.0040
