Today we will be talking about a fascinating topic, one of which seems to go under the radar from time to time and yet has an extreme impact on the populations of modern humans (us)! We will discuss bottlenecks, what they are, and how they affect the populations they affect. This is not strictly based on what happens to humans, but that, of course, will be the main point of this article. Humans.
A human population bottleneck is a sharp reduction in the size of a human population due to environmental events or human activities, such as famines, earthquakes, floods, fires, disease, droughts, wars, migrations, or cultural practices. Population bottlenecks can significantly affect the genetic diversity and health of human populations and their evolutionary history and adaptation. A bottleneck can occur when there is any reason or reasons for drastically reducing the genetic pool available to a population at any given point. By reducing the gene pool and minimizing admixture, we can begin to see the issues arise that small populations face. But of course, we survived and possibly even came out better and more adapted to our ecological niches at the end of each of these proposed bottlenecks.
Genetic diversity is the variation in the genes of a population. It is essential for the survival and adaptation of a population, as it allows for natural selection to act on beneficial traits and eliminate harmful ones. Genetic diversity also provides the raw material for innovation and creativity in human culture and technology. If there is a small gene pool, we are going to see the same phenotypic features over and over, and if those features, for whatever reason, do not fit the new niche created by the bottle neck, much of the population will die off. For one reason to another, infighting, lack of breeding opportunities, or higher competition for resources such as food and water. Population genetics is very fragile, and changes in how things are done can have drastic impacts.
Population bottlenecks can reduce genetic diversity by increasing the effects of genetic drift and inbreeding. Genetic drift is the random change in the frequency of alleles (different versions of a gene) in a population due to chance events. Inbreeding is the mating of closely related individuals, which increases the likelihood of inheriting identical copies of alleles from both parents. Both genetic drift and inbreeding can lead to the loss of alleles, the fixation of alleles (when one allele becomes the only one in a population), and the increase of harmful recessive traits that can cause diseases or impairments.
When we are dealing with smaller populations that are trying to survive, any evolutionary disadvantages they face are going to be increased. If a recessive trait is usually bred out of the population, allowing the population to flourish and grow, but only those individuals with this allele or trait remain, then whatever negative aspect about this feature will cause damage. Sometimes too much damage to the area’s ecology, and we have extinctions, or, as we are learning, near extinction events called bottlenecks.
Population bottlenecks can also affect the evolutionary history and adaptation of human populations by altering their patterns of genetic variation and relationships with other populations. For example, population bottlenecks can create founder effects, which occur when a small group of individuals colonizes a new area or becomes isolated from the main population. Founder effects can result in genetic differences between the new population and the original one and among different populations that originated from the same source. Founder effects can also influence the cultural and linguistic diversity of human populations. At its most extreme, many anthropologists, when asked the question, “Where will humans be in a thousand years?” It is hard to answer, for if we do go into space for any extended time, and I do mean a long amount of time, those who went into space would be the “founders”; setting a new population base wherever they went, one which may not exactly represent the original gene pool back at home. The founder effect has had a great role in our development and evolution; it has allowed us to develop and to develop changes, changes which make is beautifully part of the same species.
Examples of human population bottlenecks
Human populations have experienced many population bottlenecks throughout their history, some of which have left traces in their genomes that can be detected by genetic analysis. Some examples are:
– The Out-of-Africa bottleneck: This occurred when a small group of modern humans left Africa about 60,000 to 80,000 years ago and spread across the world, replacing other hominin species such as Neanderthals and Denisovans. This bottleneck reduced the genetic diversity of non-African populations compared to African ones, as well as introduced some genetic variants that were adaptive to different environments outside Africa.
– The Toba catastrophe bottleneck: This is a controversial hypothesis that proposes that a massive volcanic eruption at Lake Toba in Indonesia about 74,000 years ago caused a global cooling event that drastically reduced the human population to about 10,000 to 30,000 individuals. This bottleneck would have further decreased the genetic diversity of human populations and increased their genetic differentiation.
– The Ashkenazi Jewish bottleneck: This occurred when a small group of Jewish people migrated from Europe to Eastern Europe about 1,000 years ago and established a distinct cultural and religious identity. This bottleneck reduced the genetic diversity of Ashkenazi Jews compared to other Jewish groups and increased their susceptibility to certain genetic diseases such as Tay-Sachs disease and cystic fibrosis.
– The New Zealand black robin bottleneck: This occurred when only five individuals of this bird species survived after habitat destruction and predation by introduced mammals in the 1980s. This bottleneck severely reduced the genetic diversity and fitness of this species, which required intensive conservation efforts to prevent its extinction.
So there we have it! I hope this has answered some of your questions surrounding what bottlenecks are when we are talking about genetics, human genetics, what it means, and the effects it could have evolutionarily on a species.
In his own words, “I am a PhD candidate in the School of Human Evolution and Social Change at Arizona State University and an affiliate of the Institute of Human Origins. I received my BA in Anthropology from Stony Brook University and my MA in Anthropology from the University of Victoria. My research focuses on the emergence and role of pyrotechnology in human adaptations during the late Middle and Later Stone Age of South Africa.
I have conducted field work at the late Lower Paleolithic site Shishan Marsh – 1 in al-Azraq, Jordan and the Upper Paleolithic cave site Mughr el-Hamamah in the Jordan Valley. Currently, I am doing field work at a Later Stone Age site called Knysna in South Africa.
My hobbies include PC gaming, skateboarding, and flintknapping.”
You can contact him to get a chance to look at his SOP (Statement of Purpose) and his CV: jkmurra5@asu.edu
If you learned something, which I sure hope you did, be sure to drop a like and subscribe to see more interviews like this one and plenty of other content!
Bipedalism, or walking on two legs, is one of the defining characteristics of humans. But why do we walk this way? There are many hypotheses about the evolution of bipedalism. Still, the most likely explanation is that it allowed our early ancestors to travel long distances more efficiently, as in they could go farther while using fewer calories and explore the world even further.
While Bipedalis may not be the thing that “makes us Human,” as I believe there is no one such thing, it is an aspect of our evolutionary history and our current lives that cannot be ignored; its importance in understanding who we are is critical.
The advantages of bipedalism
Bipedalism has many advantages over quadrupedalism, or walking on four legs. For one, it allows us to travel long distances more efficiently. This is because bipedalism frees up our hands, which we can use to carry tools or food. Additionally, bipedalism allows us to see over tall grass and other obstacles, which gives us a better view of our surroundings.
Bipedalism also has some social advantages. For example, it allows us to stand face-to-face with each other, which is essential for communication and cooperation. Additionally, bipedalism allows us to use our hands for gestures and other nonverbal communication.
We can run, walk long distances, and get around places no other animals can, thanks to these strange spindly things that come off the bottom of our torsos. But being bipedal is not always a good thing. As we will soon see.
The disadvantages of bipedalism
Bipedalism also has some disadvantages. For one, it puts more stress on our legs and feet. Additionally, bipedalism makes climbing trees or running as fast as quadrupeds more difficult. But these are the simple things that make the disadvantages of bipedalism apparent.
Less apparent is the back strain, the heavyweight our bones have to move around to keep us upright. It is a delicate balancing act. As Dr. Jeremy DeSilva put it in his book First Steps; How Walking Made Us Human (Which I highly recommend), he mentions that we are always constantly in a controlled fall. We put one leg forward and fall controlled until we throw the other leg out in front of us. This is very true when you think about it, and there is so much room for error when doing this.
We fall constantly, and the older we get, the more dangerous and likely these events are to occur. Taking a tumble on the football field when you are 18 and falling on your hip when you are 80 will feel very different.
The evolution of bipedalism
The exact evolutionary history of bipedalism is still a matter of debate, but it is thought to have evolved over millions of years. We have examples of bipedalism that may date back to seven million years ago, close to our separation from the pan family. Sahelanthropus tchadensis may be the oldest hominin we know of. But it is still debated on whether or not it was bipedal, to begin with.
The next candidate would be the millennial man, or Orrorin Tungunensis, which dates to around 5-6 million years ago.
Another great example of ancient bipedalism is the Laetoli footprints in Tanzania. It contains multiple tracks of footprints made by something remarkably human, thought to be Australopithecus afarensis (save the site A trackway). We can see how these footprints were made because they were imprinted on top of recently deposited volcanic ash, so we can even reliably date them to about The Laetoli footprints are a series of hominin footprints that were discovered in Laetoli, Tanzania, in 1978. The footprints are dated to 3.6 million years ago, and they are the oldest known direct evidence of human or human-like bipedalism.
The footprints were discovered by Mary Leakey and her team during an archaeological excavation at Laetoli. The footprints were found in a layer of volcanic ash that had been laid down by a volcano that erupted about 3.6 million years ago. The ash layer had preserved the footprints in excellent condition.
The footprints are about 1.7 meters long and 0.7 meters wide. They show that the hominin who made them was walking upright, with its feet flat on the ground. The footprints also show that the hominin was walking at a slow pace.
The Laetoli footprints are a significant discovery because they provide direct evidence of human or human-like bipedalism at a time when it is thought that hominins were still evolving from ape-like ancestors. The footprints also provide evidence that hominins were walking upright in East Africa at least 3.6 million years ago.
The Laetoli footprints are displayed at the National Museum of Natural History in Washington, D.C.
Conclusion
Bipedalism is a complex trait with many advantages and disadvantages. It is thought to have evolved over millions of years, and it has played a major role in the evolution of humans.
References
Day, M. H. (1986). Hominid footprints from Laetoli, Tanzania. Nature, 322(6081), 17-21.
Lovejoy, C. O. (1981). The evolution of human walking. Scientific American, 244(5), 118-125.
Tuttle, R. H. (1987). The primate origins of human bipedalism. New York: Columbia University Press.
Science communication is the process of sharing scientific information with the public. It can be done through various channels, including news media, social media, public talks, and educational materials.
Science communication is essential for several reasons. First, it helps to build public understanding of science. People can better make informed decisions about their lives when they understand science. For example, if people understand the science of climate change, they are more likely to support policies that address it.
Second, science communication can help to increase public trust in science. People who trust science are more likely to support scientific research and innovation. This is important because scientific research is essential for solving many of the world’s most pressing problems.
Third, science communication can promote scientific literacy. Scientific literacy is understanding and using scientific information to make decisions. When people are scientifically literate, they can better participate in public debates about science and technology.
There are many challenges to science communication. One challenge is that science can be complex and difficult to understand. Another challenge is that the public often has negative perceptions of science. This can be due to some factors, such as the media’s portrayal of science or the public’s lack of understanding of scientific methods.
Despite these challenges, science communication is essential for a healthy democracy. When the public understands science, they can better make informed decisions about their lives and support policies that promote scientific research and innovation.
There are several ways to improve science communication. One way is to make scientific information more accessible to the public. This can be done by using plain language, avoiding jargon, and providing visuals and examples. Another way to improve science communication is to engage with the public in a two-way dialogue. This means listening to the public’s concerns and questions and providing them with opportunities to participate in scientific research.
Science communication is an integral part of the scientific process. It helps to ensure that scientific research is conducted in a way that is transparent and accountable to the public. It also helps to ensure that the public has the information they need to make informed decisions about their lives.
References
National Academies of Sciences, Engineering, and Medicine. (2017). Communicating science effectively: A research-based guide for scientists and engineers. Washington, DC: The National Academies Press.
National Science Foundation. (2018). Science and technology in the public sphere: A research agenda for science communication. Arlington, VA: National Science Foundation.
Royal Society. (2012). Communicating science effectively: A guide for researchers. London: The Royal Society.
Hello everyone, it’s great to have you back on my channel! Today, I am excited to introduce you to our special guest, Genevieve von Petzinger. She is a renowned paleoanthropologist and the author of the captivating book, “First Signs: How Ancient Symbols Shaped Humanity”. In this book, she delves into the fascinating origins and meanings of the geometric symbols that were created and painted on cave walls by our ancestors across the globe. Genevieve also shares her amazing journey of exploration to some of the most remote and intriguing sites on Earth. During our conversation, we discussed her book, her research, and the valuable insights that these ancient symbols can offer us about our past and ourselves. I hope you found this interview as intriguing as I did, and please remember to like, comment, and subscribe for more fascinating content. Thank you for tuning in!
Hi everyone! Welcome to The World of Paleoanthropology, where I share my passion for human evolution and archaeology. Today I want to talk about the earliest stone tool industries, from the Lomekwian to the advanced Homo sapiens tools. How did our ancestors and relatives develop such amazing skills and technologies? What are lithics, and how are they made? Let’s find out in this very brief overview!
Lithics is a term that refers to stone tools or any other objects made from stone. Lithics can be classified into different types based on how they are made, such as flaked, ground, and polished stone tools. Flaked stone tools are the most common type of lithics, and they are made by striking a piece of stone (called a core) with another stone (called a hammerstone) or a bone to detach sharp flakes that can be used for cutting, scraping, piercing, etc.
The oldest known flaked stone tools are from the Lomekwian industry, named after the site of Lomekwi 3 in Kenya, where they were discovered in 2015. These tools date back to 3.3 million years ago, predating the genus Homo by 700,000 years! That means that some of our earlier ancestors or relatives, such as Australopithecus afarensis (Lucy’s species), could make and use stone tools . The Lomekwian tools are very simple and crude, consisting of cores, flakes, hammers, and anvils. They were probably used for breaking nuts or bones to extract the marrow.
The next major stone tool industry is the Oldowan, named after the site of Olduvai Gorge in Tanzania, where it was first discovered in 1964. The Oldowan tools date from 2.6 to 1.7 million years ago, and they are associated with the earliest members of the genus Homo, such as Homo habilis (the “handy man”) . The Oldowan tools are more refined and diverse than the Lomekwian ones, including choppers with one sharp edge, scrapers, awls, etc. They were used for processing meat, plants and other materials.
The Oldowan industry was followed by the Acheulean industry, named after the site of Saint-Acheul in France, where it was first discovered in 1859. The Acheulean tools date from 1.7 million to 100,000 years ago, and they are associated with Homo erectus (the “upright man”) and later Homo species. The Acheulean tools are more complex and symmetrical than the Oldowan ones, and they include handaxes with two sharp edges, cleavers, picks, etc. They were used for hunting, butchering, woodworking and other tasks .
The Acheulean industry was followed by several regional industries that developed in different parts of the world during the Middle Paleolithic (300,000 to 50,000 years ago) and the Upper Paleolithic (50,000 to 10,000 years ago) periods. These industries include the Mousterian (associated with Neanderthals), the Aurignacian (associated with modern humans), the Solutrean (known for its leaf-shaped points), the Magdalenian (known for its bone and antler tools), etc. These industries show a great diversity and sophistication of lithic technologies, such as blade production, pressure flaking, retouching, hafting, etc. They also show evidence of symbolic behavior and artistry, such as engraving, painting and sculpting on stone and other materials.
As you can see, the history of stone tool industries is a fascinating story of human evolution and innovation. From the simple Lomekwian tools to the advanced Homo sapiens tools, our ancestors and relatives have demonstrated remarkable cognitive abilities and cultural adaptations. I hope you enjoyed this blog post and learned something new. Stay tuned for more posts on human evolution and archaeology!
In the scientific field of paleoanthropology, and paleontology overall, being able to deduce how an organism moved is very useful. This is because it can show how an organism lived and can give insight into its evolutionary history. Most of the time, all paleontologists have are fossils, but this can be surprisingly helpful. For paleoanthropologists specifically, being able to do this is crucial, as it can help determine how a hominin is related to other species and therefore, its place in our family tree.
The field concerned with determining how an organism moved/moves is known as biomechanics. Biomechanics is the study of the structure and function of biological systems. When combined with paleontology, biomechanics can give great insight into how an extinct animal moved, pulling from an array of morphological characteristics. Biomechanic scientists look for and examine all these characteristics in fossils to get a good idea of how extinct organisms moved.
Animals move in all sorts of locomotor styles. Fish often move by slowly moving their tails back and forth to propel themselves through the water. Most land animals move quadrupedally (on four legs), and retain a pronograde posture. This means that their bodies are more horizontally positioned. Many animals that walk this way have sprawled out limbs, such as in lizards, whereas others have limbs tucked beneath them, such as in quadrupedal mammals like cats and dogs.
However, many animals walk bipedally (on two legs). Many theropod dinosaurs walked this way, including Tyrannosaurus rex and even modern birds. Some mammals walk this way too, such as kangaroos. This is the locomotor style of humans. There is one thing that makes us different from other bipedal animals however, and that is our posture. Humans retain an orthograde posture, rather than a pronograde posture. This means that our posture is upright/vertical. Some other primates, such as gibbons, possess an orthograde posture as well. Most of the time, gibbons move via suspensory brachiation, where they swing from branch to branch with their front arms. Gibbons also can move bipedally when need be, but it is not their main method of locomotion.
The first known hominin that possesses these locomotor styles is called Sahelanthropus tchadensis, which lived roughly 7 million years ago. Sahelanthropus, along with all the other fossil hominins, possess many different traits that allow for them to walk bipedally. These traits can be found from the head to the feet. Most are directly related to the animals’ locomotion, though some are indirectly related. Some even have other side effects not related to locomotion as a result. Lets go over all these traits from top to bottom:
The Skull:
The first trait is found in the skull. More specifically in the basicranium (the lower part of the skull). This trait has to do with the foramen magnum, the hole at the base of the skull where the spinal cord enters. In organisms that walk quadrupedally, their foramen magnum is more posteriorly positioned, meaning it’s towards the back of the skull. This allows for the spinal cord to enter more horizontally, giving the animal a more pronograde posture. The foramen magnum is also slightly angled. Depending on the locomtion of the animal, the hole will be angled back or forward slightly. The position tells you the posture and the angle tells you the locomotion.
In bipedal species with an orthograde posture, the foramen magnum is more anteriorly positioned, meaning it is closer to the front, and is angled forward slightly. The occipital condyles, small bony projections on each side of the foramen magnum are angled as well, allowing for articulation with the cervical vertebrae. Sahelanthropus possesses this trait, though to a lesser extent than modern humans.
Diagram comparing the foramen magnum of a chimpanzee, modern human, and Sahelanthropus (Nature, 2013).
The Rest of the Body:
The next trait is the curvature of the vertebrae. In most apes, their vertebral column forms a curved ‘c’ shape. This allows them to walk on four legs. In humans however, our vertebral column forms a more ‘s’ shape. This condition in humans is known as lumbar lordosis. This curvature in the lumbar (lowermost) vertebrae places the center of mass of the body directly over the hips and legs, and distributes weight throughout the body. This makes it easier to maintain an orthograde posture and bipedal locomotion.
A comparison of the vertebral column of an extant great ape and a modern human (The Australian Museum, 2020)
This condition first appeared around 4 million years ago, with the genus Australopithecus.There are several specimens of Australopithecus that have well preserved vertebrae showing they had this human-like condition. A specimen of a juvenile Australopithecus afarensis dating to 3.3 million years ago shows a very human-like condition in the vertebrae. The specimen, DIK-1-1, was nicknamed “Selam” after the Amharic word for peace, or the “Dikika child”, after the place she was found. Selam was a very significant discovery, and showed lots about the evolution of human growth patterns, but most importantly, she had a fully articulated human-like vertebral column.
The preserved vertebrae and scapula of DIK-1-1 showing the human-like condition (UChicagoMedicine, 2017).
Though lumbar lordosis is beneficial when it comes to bipedalism, it doesn’t come without cost. Lumbar lordosis can cause severe pain and result in several medical problems, such as DMD (Duchenne muscular dystrophy) and walking difficulties.
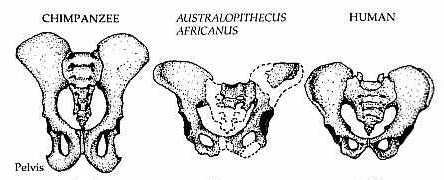
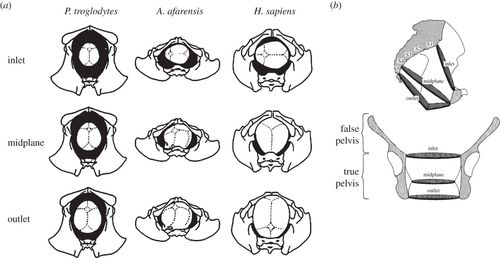
Lower down in the body, we find perhaps the most important trait: The orientation of the pelvis. In bipedal species, the iliac crests (the upper, wing-like part of the hip) are sagittally oriented. This means they are oriented into the midline of the body. This gives the pelvis a more bowl-like shape. This is further supported by a wider sacrum bone than in other apes. This allows us to balance upright without having to shift our weight forward, which would require us to be quadrupedal. It also plays a role in thermoregulation, as it lowers the surface area-to-mass ratio, allowing for more heat loss.
Diagram comparing the pelvises of a chimpanzee, Australopithecus africanus, and a modern human (The Haps Blog, 2015)
This evolutionary advantage comes with a trade off, an evolutionary compromise, however. With the narrow bowl-like pelvis, giving birth is much more difficult. In humans, the baby’s needs to rotate 3 times to fit through the birth canal, prolonging the average birth time to 14 hours, compared to other great apes, such as chimpanzees, which have an average birth time of 2 hours. Another reason why other apes have it so easy is because of the small heads of the babies being born. The ability to walk bipedally outweighed the need for short births, so we compromised one trait over another.
An inferior view of the birth canal of a chimpanzee, Australopithecus afarensis, and a modern human, showing the head rotation of the child during birth (National Library of Medicine, 2015).
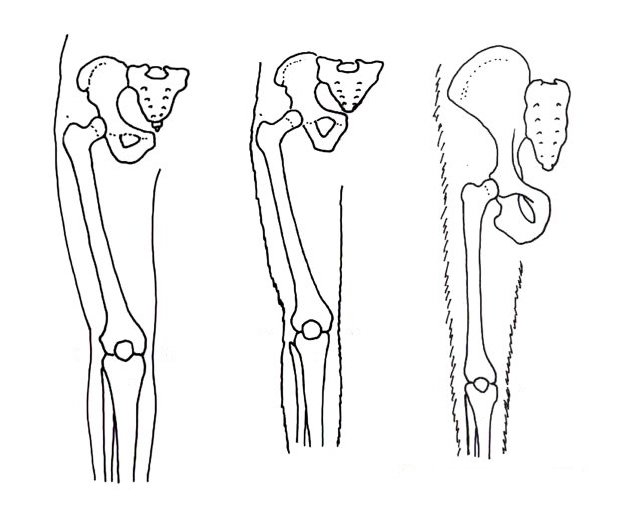
Below the pelvis, the femora (bones in the thigh) can also give a good clue of how a hominin walked. Organisms that walk bipedally possess a bycondylar angle, a slight angle in the femur that places the individual’s body mass beneath them, which is advantageous for this method of locomotion. Along with this, the epiphysis (the head of the femur) is thicker than in quadrupedal apes. This places the knee in a valgus (angled away from the midline) position. The condyles at the end of the femur are also thicker, allowing them to bear more weight while walking.
A diagram comparing the bycondylar angle in a modern human, an extinct hominin, and chimpanzee (The Australian Museum, 2020).
The oldest known hominin that verifiably possesses this trait is Orrorin tugenensis, an early hominin which lived in Kenya 6 million years ago. Orrorin is known from little fossil material, but what is known of it reveals lots about its locomotion and evolution. The most complete femur from this species is known as BAR 1002’00. This specimen is a left femur, and possesses an obvious bycondylar angle. Along with this, BAR 1002’00 has a thick epiphysis, meaning that Orrorin had the capability of bipedal locomotion, though it may not have been habitual.
Interestingly, the thickness of the epiphysis and the bycondylar angle are to a greater extent than what is found in later Australopithecus. This suggests that Orrorin was an off-branch species that evolved more bipedal traits before the rest of the hominins.
The BAR 002’00 femoral specimen of Orrorin tugenensis (Stony Brook University, 2013).
Right beneath the femur, the knee can also give a clue about how an organism walked. Human knee joints can fully extend and lock, allowing for the leg to extend straighter out. This trait is less significant, and it is unknown when it first appeared, but it could have come about anywhere from 4-2 million years ago.
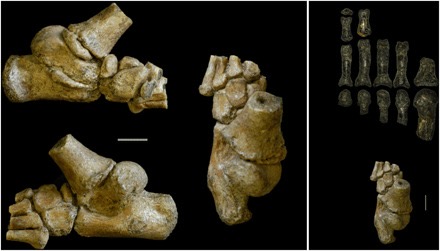
At the very bottom of the body, the feet have several traits that allow for bipedal locomotion. Firstly, is the hallux bone. The hallux (big toe) in most primates is divergent, forming a more thumb-like shape. This makes the foot resemble a hand, allowing for the animal to be able to grasp branches with its hands and feet. Some hominins, such as Ardipithecus, possessed this trait, suggesting they were still spending time in the trees 4.4 million years ago, though the foramen magnum position suggests this genus still walked bipedally.
Cladogram showing the divergent hallux throughout the superfamily Hominoidea, proposing two different hypotheses for the evolution of human bipedalism (eLife, 2019).
The divergent hallux isn’t lost until 4 million years ago with the genus Australopithecus, when hominins began using terrestrial bipedalism. This shows that it was a very quick change.
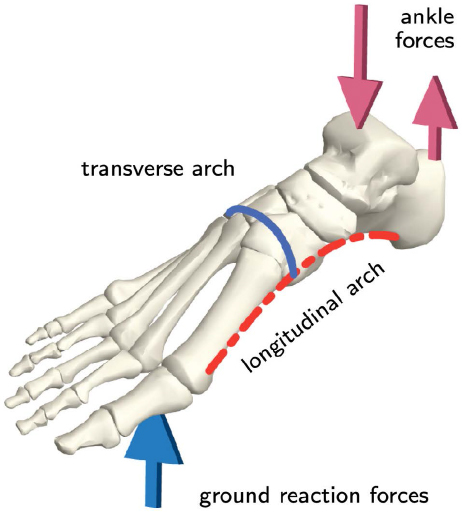
Aside from the hallux, the arches in the foot are also important for bipedal locomotion. Humans have three arches in our feet, the medial and lateral longitudinal arches, and the anterior transverse arch, while other apes have none. The transverse arch makes the foot stiffer, allowing it to bear more weight. This is useful for propulsion, allowing humans to walk bipedally more efficiently. The transverse arch specifically is significant as it contributes over 40% of the foot’s stiffness.
Diagram of the human foot showing the transverse and longitudinal arches, and the forces placed on the foot (Anatomy and Biological Anthropology, 2021).
Hominin fossil feet are very rare, but there are several fossil specimens of hominin feet showing these arches. The first group to fully possess this trait are the Australopiths. Selam, the juvenile Australopithecus specimen mentioned earlier, has a surprisingly well preserved foot. The calcaneus (heel bone) of Selam is very well preserved and is very human-like. The heel morphologies place the tibia (shin bone) orthogonally (vertically over) foot as in modern humans, but the growth rate of the foot resembles more of other great apes. Other fossils of adult individuals of Australopithecus afarensis show morphologies more similar to adult humans.
The foot specimen of DIK-1-1 (National Library of Medicine, 2018).
Another foot specimen, this time from Homo habilis, an early member of our genus from 2.5 million years ago shows similar, human-like morphology. The specimen, known as 0H 8, belonged to an adult individual, and is very similar to that of modern humans. The robustness of the foot and the foot arches are very similar to ours. These two finds show that from 4-2 million years ago, the foot morphology of hominins became much more adapted for terrestrial bipedalism and started resembling that of modern humans very quickly.
Now that we’ve looked at all the traits directly tied to bipedalism, let’s look at one which is indirectly related.
Because our tribe, Hominini, has specific traits, such as bipedal locomotion, if we find one fossil with other traits associated with hominins, it is reasonable to assume that they walked bipedally. Why would it have some traits but not the others? This isn’t the most reliable method, as evolution can often be very unpredictable, but it can be useful if it is all you have, and often times, it is all you have.
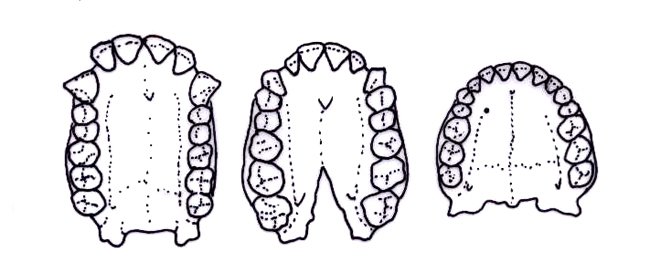
One of these traits is the size of the maxillary (upper) canines. In most apes, and primates in general, individuals have very large canines. These canines are often used for sexual and warning displays. Oftentimes for primates, the bigger the canines the better. The size of the canines also allows for a honing complex, where the upper canines are sharpened against the first premolars. This also resulted in the absence of a diastema, a gap in between the lower incisors and canines and upper canines that allowed for space for the large upper canine.
Diagram of the upper jaws of a chimpanzee, Australopithecus, and modern human, showing the canine size and diastema (The Australian Museum, 2018).
As hominins began evolving bipedal locomotion, their maxillary canines began being reduced. This allowed for non-honing chewing, giving our jaws more movement. Ardipithecus ramidus is the first species that shows significant canine reduction, and also has very little sexual dimorphism. It is around this time from 7-4 million years ago that hominins lost their large canines and began evolving bipedalism, so if a fossil ape maxilla is found with reduced canines, it very well could be a bipedal hominin, though you should also have precautions and never jump to conclusions.
A diagram showing the canines of a chimpanzee and Ardipithecus kadabba (UCBerkleynews, 2004).
There are other ways to tel how a species walked without physical fossil remains. Those are trace fossils, or footprints.
There are several preserved trackways from hominins showing their foot morphology and how they walked. The most famous of these trackways seems to be the Laetoli footprints from Tanzania.
Found in Tanzania, in a site known as Site G, roughly 70 hominin footprints are known, spanning about 88 feet. These tracks date to about 3.6 million years ago, and were likely made by 2, possibly 3 individuals of Australopithecus afarensis who were walking bipedally along a wet volcanic ashfall.
These tracks show a foot morphology very similar to modern humans, and a heel-strike method of walking, in which the heel touches the ground first, just like n modern humans, though 1.5 million year old tracks in Koobi Fora, Kenya, produced by Homo ergaster show an even more modern-like foot morphology and weight transfer, making the Laetoli footprints more primitive.
Research suggests that the hominins which produced the Laetoli footprints had stride lengths similar to modern humans, but there is debate on whether they used a bent-hip, bent-knee gait, similar to a chimpanzee, or an extended-hindlimb gait, similar to a modern human. This debate still seems to be unresolved, but there is lots of research going into it.
The Laetoli Footprints (Science News, 1976).
Conclusion:
After examining all the morphological characteristics necessary for bipedal locomotion, it is very easy to conclude how a hominin walked only from fossil remains. There are many traits, from the head to toes, many of which are insignificant or unexpected, but important nevertheless. Even without physical fossils, footprints produced by many different hominin species also can give a good idea on how these animals walked. It is important to give a good and honest look, as no matter what you have, it could very well give insight on the lifestyle, behavior, and evolution of our ancient ancestors.
Williams, A. S., Zeng, I., Paton, J. G., Yelverton, C., Dunham, C., Ostrofsky, R. K., Shukman, S., Avilez, V. M., Eyre, J., Loewen, T., Prang, C. T., Meyer, R. M. (2022). Inferring lumbar lordosis in Neandertals and other hominins. PNAS Nexus, 1(1), pga005. https://doi.org/10.1093/pnasnexus/pgab005
Filiz, B. M., Toraman, F. N., Kutluk, G. M., Filiz, S., Doğan, K. S., Çakir, T., Yaman, A. (2021). Effects of lumbar lordosis increment on gait deteriorations in ambulant boys with Duchenne Muscular Dystrophy: A cross-sectional study. Brazilian Journal of Physical Therapy, 25(6), 749-755. 10.1016/j.bjpt.2021.05.001
Gruss, T. L., Schmitt, D. (2015). The evolution of the human pelvis: changing adaptations to bipedalism, obstetrics and thermoregulation. Philosophical Transactions B, 370(1663), 20140063. 10.1098/rstb.2014.0063
Almécija, S., Tallman, M., Alba, M. D., Pina, M., Solà-Moyà, S., Jungers L. W. (2013). The femur of Orrorin tugenensis exhibits morphometric affinities with both Miocene apes and later hominins. Nature Communications, 4, 2888. https://doi.org/10.1038/ncomms3888
Pickford, M., Senut, B., Gommery, D., Treil, J. (2002). Bipedalism in Orrorin tugenensis revealed by its femora. Comptes Rendus Palevol, 1(4), 191-203. https://doi.org/10.1016/S1631-0683(02)00028-3
Harcourt-Smith, E. H. W. (2010). The First Hominins and the Origins of Bipedalism. Evolution: Education and Outreach, 3, 333-340. https://doi.org/10.1007/s12052-010-0257-6
Prang, C. T. (2019). The African ape-like foot of Ardipithecus ramidus and its implications for the origins of bipedalism. eLife, 8, e44433. https://doi.org/10.7554/eLife.44433
Asghar, A.l, Naaz, S. (2021). The Role of Transverse Arch in Foot Stiffness and Its Clinical Implications. Anatomy and Biological Anthropology, 34(3), 103-104. https://doi.org/10.11637/aba.2021.34.3.103
DeSilva, M. J., Gill., M. C., Prang, C. T., Bredella, A. M., Alemseged, Z. (2018). A nearly complete foot from Dikika, Ethiopia and its implications for the ontogeny and function of Australopithecus afarensis. 4(7), eaar7723. 10.1126/sciadv.aar7723
DeSilva, M. J., Zipfel, B., Arsdale, V. P. A., Tocheri, W. M. (2010). The Olduvai Hominid 8 foot: Adult or subadult? Journal of Human Evolution, 58(5), 418-423. https://doi.org/10.1016/j.jhevol.2010.03.004
Delezene, K. L. (2015). Modularity of the anthropoid dentition: Implications for the evolution of the hominin canine honing complex. Journal of Human Evolution, 86, 1-12. https://doi.org/10.1016/j.jhevol.2015.07.001
Suwa, G., Sasaki, T., Semaw, S., White, D. T. (2021). Canine sexual dimorphism in Ardipithecus ramidus was nearly human-like. PNAS, 118(49), e2116630118. https://doi.org/10.1073/pnas.2116630118
Raichlen, A. D., Gordon, D. A., Harcourt-Smith, H. E. W., Foster, D. A., Haas Jr., R. W. (2010). Laetoli Footprints Preserve Earliest Direct Evidence of Human-Like Bipedal Biomechanics. PLoS ONE, https://doi.org/10.1371/journal.pone.0009769
Raichlen, A. D., Pontzer, H., Sockol, D. M. (2008). The Laetoli footprints and early hominin locomotor kinematics. Journal of Human Evolution, 54(1), 112-117. https://doi.org/10.1016/j.jhevol.2007.07.005
Now that I have had some time to sit down and relax after getting home from almost two weeks of traveling between the Bill Kimbel inaugural Lecture at the new IHO campus in Tempe, Az, and then, of course, to the AABAs in Reno, Nv, I thought it would be fun to tell you, folks, about how it went, what I learned about my first conference, and what I can Impart onto any future conference goers, as well as give a window into what was going on in general! It was organized chaos! But it was terrific, and I have very few if any, negative things to say at all.
So let’s get into it!
We will go in chronological order; thus, we will start with Dr. Bill Kimbel Inaugural Lecture, the first in the new lecture series put on by the Institute of Human Origins (IHO) and Arizona State University (ASU), which was on April 15th, a Friday. We arrived that afternoon at our hotel, got ready, and proceeded to the new Tempe ASU campus for the cocktail hour and lecture, which was going to be given by famed Dr. Bernard Wood, a close friend of Dr. Kimble’s.
I will be candid this was the first event of its kind that I have attended in my adult life, I had been to plenty of fancy events as a child for my parent’s work, but none that I was attending for my wants, so it was a little awkward. I did not know anyone save Kota, my partner of nearly ten years. We tootled around a little, ate some the good food, and looked at the silent auction hosted to benefit a new undergraduate position at ASU to continue Bill’s work; at least, that is the hope.
We all got seated in the theatre for the lecture, which was a lovely venue on the beautiful newly built Tempe campus. The lecture was great, there were funny and sad parts, and overall it was very educational, learning about the work Dr. Kimble had touched and the many ways he affected the field. His family was there, and the whole event was a great experience. I tried to remember how solemn of an event it was while also enjoying something wholly new to me – a part of the field that I had never seen. I would find myself thrust into a portion of the field in less than a week!
After the lecture, we returned home for a few days before heading to Reno for the 92nd annual AABA conference, which would also be the first event I would attend; I was thrilled! I was excited to go and meet so many people I had met online and interviewed on my Youtube show “The Story of Us,” as I knew many of my guests would be in attendance.
Aside from a flat tire on the morning, we were to depart and a delayed dinner, the drive from Los Angeles to Reno went without a problem, and we arrived the evening before the conference started.
The first day I attended a workshop on understanding how to analyze ancient DNA data was fun. We got to do some of the work on our computers (for those who could get it working), and that was very educational and a skill I will need to practice, but I now know how to do it! After that, I met with the first people I knew, chatted for a bit, and then explored until it was time for dinner, which I had with the beautiful Dr. Briana Pobiner, who had been helping me prepare for the conference. Still, I would soon find that would take me under her wing for the entirety of the week! I could not have been more fortunate to have someone like that in my corner!
After dinner, it was time for the opening reception, which I quietly snuck into; the introvert that I am, unable to hide from Briana for long before she was so kind as to introduce me to practically everyone she knew who walked in the door. Talk about making connections!
The next few days were filled with awesome symposiums, including an entire day dedicated to the work Dr. Bill Kimbel affected in the field, with many famous individuals showing off the result they were involved in with Bill. Some of the other highlights were, surprise! Briana would shock me again by inviting me as her +1 to the President’s Dinner! Talk about imposter syndrome! I was having trouble being there in general, but now to go to this exclusive event, where I chatted, drank, and met even more people! It was terrific, throwing another curveball at me; after the night, Dr. Pobiner invited me to the education committee meeting, which I imagined consisted of quite a few people, and I would slide in unnoticed.
The next day, my alarm went off an hour late, and I awkwardly walked into a small room with a long table and about 10-15 people sitting around it! This was more of a board meeting than I was expecting! I sat quietly, respecting my current position, and learned a great deal about where the AABA is going with its outreach in the coming year; I learned some things I could apply to you guys and even gave a tip or two! It was great!
After another day of biological anthropology fun, it was finally time for the closing reception and to say our goodbyes to everyone, but not before meeting even more new people, scheduling some episodes of “The Story of Us,” and just having a great time! I love being among my people for the first time that I can truly remember, feeling like I belonged in a way that I have not before, not having to explain my thoughts or ideas, but having people already know of them, or have ideas of their own! Being among like minded people was so utterly refreshing!
Leaving the following day, we had a safe and sound drive home, and I cannot wait and am already planning for the AABAs 2024, which will be in my backyard in Los Angeles!
Welcome to the World of Paleoanthropology, where we explore the origins and evolution of our human ancestors. In this video, we will introduce you to one of the most fascinating and mysterious discoveries in recent years: the skull and skeleton of Neo, a member of the Homo naledi species.
Homo naledi is a new species of ancient human relative that was first announced in 2013. It has a mix of primitive and modern features, such as a small brain like an early human, but hands and feet like a modern human. It also has a pelvis and shoulders like those of an ape-like Australopithecus.
Neo is the most complete skeleton of Homo naledi ever found. It was discovered in 2017 in a remote chamber of the Rising Star cave system in South Africa, along with two other individuals. Neo was an adult male who stood about 1.4 meters tall and weighed about 40 kilograms. He lived between 236,000 and 335,000 years ago, which means he may have overlapped with our own species, Homo sapiens, in Africa.
In this video, we will show you how Neo’s skull and skeleton reveal new insights into the anatomy, behavior, and culture of Homo naledi. We will also explore some of the mysteries and controversies surrounding this species, such as how they got into the cave, why they buried their dead, and what their relationship was with other ancient humans.
Join us as we journey into the past and meet Neo, one of our long-lost cousins in the human family tree. If you enjoy this video, please like, share, and subscribe to our channel for more paleoanthropology content.