**Season 2 Finale: “Unveiling the Mysteries: The Latest in Paleoanthropology Discoveries”** https://podcasters.spotify.com/pod/show/world-of-paleoanthropolog/episodes/Season-2-Finale-Unveiling-the-Mysteries-The-Latest-in-Paleoanthropology-Discoveries-e2k848a Available on Apple Podcasts and soon the Video Version on YouTube! Enjoy! See you for Season Three!!!
Snake Detection Theory: Did Snakes Play a Role in Primate Evolution?

Just like primates, snakes are one of the most unique vertebrates on the planet. They have evolved to adapt for various environmental conditions, occupying various niches across ecosystems, across the entire world. Ophidiophobia, the psychological fear of snakes, is one of the most common fears that humans seem to innately develop, often without ever encountering a snake. It’s believed that common phobias, especially those dealing with real objects, may stem from a deeply-ingrained evolutionary instinct.
Written by — Gabriel Stroup
Snake Detection Theory (SDT), an overarching hypothesis within evolutionary theory, consisting of numerous sub-hypotheses, posits that many primate characteristics came about as results of interactions with predatory snakes, among other evolutionary pressures. Anthropologist Lynne Isbell (2006) has put forward a detailed, hypothetical timeline of evolutionary changes that may have occurred to the anthropoid clade of mammals since their inception in the Cretaceous, highlighting the role of snakes as predators or otherwise sources of danger, which may have subsequently influenced the evolution of primates (Figure 1).

Adapted from Isbelle, 2006.
In primatology, SDT is supported by many studies that investigate non-human primate reactions to different snake species; moor macaques, for example, have been shown to discriminate local and dangerous snakes from non-local and non-dangerous snakes, as well as to discriminate constrictors from venomous snakes (Clara et al, 2021).

Moreover, psychological studies have shown that humans have a particular talent for picking out snake silhouettes much more quickly than any other potentially-dangerous animal (Kawai, 2016). In this experiment, researchers tested a handful of undergraduate students by presenting images of various animals that are initially obscured by random white noise, but then gradually revealed through 20 steps (see Figure 2). The results show that most participants were able to correctly identify the snake between steps 6 and 9, while most other animals were not identified until steps 10 and later.
Other papers since Isbell’s publication have largely shown support for SDT. In cultural anthropology, some of the oldest human cultures still continuing on the planet, like the various Agta people from the Philippines, developed ways of detecting and preventing deaths from snakes, particularly from reticulated pythons, the longest species of snake in the world (Headland and Greene, 2011). These large constrictors most resemble the earliest snakes that the earliest primates would have made contact within prehistory. On a broader scale, snakes have long been feared or revered as mythological symbols for various ideas throughout many independent human cultures through time.
One neurological study has shown that human infants have an innate brain response to snake-specific stimuli, even when compared to snake-like caterpillars, which would be consistent with an evolutionarily-ingrained instinct to quickly identify a potential predator of primates (Bartels et al, 2020). Another study performed three different eye-tracking experiments, all of which cumulatively suggest that, compared to spiders, snakes are quite easily identified in challenging visual conditions (Saores et al, 2014).
Snake detection also manifests in being able to identify snake scales, which are remarkably unique in the animal kingdom. One study has shown that one part of the human brain, which is associated with emotional responses to stimuli, becomes vastly active when confronted with patterns resembling snake scales, compared to patterns that resemble lizard scales or bird plumage (Van Strien & Isbell, 2017. See Figure 3).

Perhaps due to the influence that snakes have potentially exerted onto the physiology of primates, there is evidence to suggest that snakes are evolving to survive against preemptive attacks by primates. Harris et al (2021) has provided reason to suspect that the advent of cobra venom was caused by the deadly interactions between cobras and Afro-asian primates; primarily from the fact that these primates bear resistance to cobra venom, where prosimians have not. This is in spite of the fact that cobra venom had previously evolved through at least three separate lineages of cobra.
The delivery of cobra venom, evolving as a way for the snake to defend itself from a distance, is itself possibly a result of primates that initially interacted with the snake from a distance. If accurate, these points would be more solid evidence to suggest that snakes and primates have gone through (and continue to go through) a unique coevolutionary process.
Despite the mounting support for SDT, there are still some valid questions that have yet to be thoroughly explored. Coelho et al (2019) have put forward some fair criticisms and questions towards the theory. Some of these criticisms/questions have been addressed by later research, but some still remain. Some examples are:
- Numerous animal groups evolved different ways to create and deliver venom, which are not unique to snakes, so it would make sense for primates to evolve a generalized pathway to detect such species, because evolving pathways for each individual group would be biologically expensive and impractical.
- The selective habituation hypothesis posits that prey animals begin with a general image of a predator, seemingly sensitive to many potential predator-stimuli, but then acclimate/habituate to their specific surroundings, learning to distinguish harmless environmental clues from useful or harmful clues. This would render it unnecessary for primates to evolve ways to detect snakes in particular, and instead have primates learn to recognize all threats local to their particular area.
It is also worth mentioning that despite the prevalence of ophidiophobia, there is also a large fraction of primates (humans specifically) that find joy from finding and interacting with snakes, and such joy occurs across human cultures just as much as fear occurs. Landová et al (2018) surveyed college students in Azerbaijan and the Czech Republic, about their attitudes and perceptions on various snake species presented to them. They found that both groups agreed that the most fear-inducing species (vipers in particular) were also the most beautiful. These results occurred even when the Azerbaijani group harbored a more negative attitude on snakes compared to the Czech group, and despite both groups having similar educational backgrounds (biological sciences). The researchers conclude that there must be both a generalized fear and generalized joy of snakes across socio-political boundaries (see Figure 4).

Snake Detection Theory is a fascinating idea which suggests that the coevolutionary relationship between primates and snakes stretches far back into early mammal times. SDT helps to explain a very common phobia, the innately-primate ability to distinguish snakes from other species in the animal kingdom, and other quirks of the primate experience. Future studies will be necessary before SDT becomes fully accepted, but as of now, it has much reason to be taken seriously. As a snake-keeper myself, even I get short flashes of fear when I see venomous snakes online, or even when I watch my own snakes explore their environment. While I encourage everyone to be educated on snake biology and behavior, I also understand that their fear of snakes may be a product of millions of years of primate evolution, which may have contributed to our very success as a species.
Bibliography
Bertels, J., Bourguignon, M., de Heering, A. et al. Snakes elicit specific neural responses in the human infant brain. Sci Rep 10, 7443 (2020). https://doi.org/10.1038/s41598-020-63619-y
Harris, R.J., Nekaris, K.AI. & Fry, B.G. Monkeying around with venom: an increased resistance to α-neurotoxins supports an evolutionary arms race between Afro-Asian primates and sympatric cobras. BMC Biol 19, 253 (2021). https://doi.org/10.1186/s12915-021-01195-x
Headland, Thomas N., and Harry W. Greene. “Hunter–gatherers and other primates as prey, predators, and competitors of snakes.” Proceedings of the National Academy of Sciences 108.52 (2011): E1470-E1474.
Hernández Tienda, Clara, et al. “Reaction to snakes in wild moor macaques (Macaca maura).” International Journal of Primatology 42 (2021): 528-532.
Isbell, Lynne A. “Snakes as agents of evolutionary change in primate brains.” Journal of human evolution 51.1 (2006): 1-35.
Kawai, Nobuyuki, and Hongshen He. “Breaking snake camouflage: Humans detect snakes more accurately than other animals under less discernible visual conditions.” PLoS One 11.10 (2016): e0164342.
Landová, Eva, et al. “Association between fear and beauty evaluation of snakes: cross-cultural findings.” Frontiers in psychology 9 (2018): 333.
Soares, Sandra C., et al. “The hidden snake in the grass: superior detection of snakes in challenging attentional conditions.” PLoS one 9.12 (2014): e114724.
Van Strien, Jan W., and Lynne A. Isbell. “Snake scales, partial exposure, and the Snake Detection Theory: A human event-related potentials study.” Scientific Reports 7.1 (2017): 46331.
Cover image: (C) Gabriel Stroup, GigabyteSpyder Photography
The Future of Paleoanthropology: How AI Will Revolutionize the Field in the Next Decade
The world of paleoanthropology, the study of ancient humans and their relatives, is poised for a transformative decade. Artificial Intelligence (AI) is set to revolutionize this field, providing researchers with unprecedented tools to uncover the secrets of our ancestors. In the next ten years, AI will not only accelerate discoveries but also refine our understanding of human evolution in ways previously unimaginable. This blog post explores how AI will impact paleoanthropology, from data analysis and fossil reconstruction to educational outreach and ethical considerations.

## Introduction: The Intersection of AI and Paleoanthropology
Paleoanthropology has traditionally relied on meticulous fieldwork and manual analysis of fossils and artifacts. However, the integration of AI promises to enhance these methods, enabling more efficient data processing and offering new insights into our evolutionary past. As AI technology continues to advance, its applications in paleoanthropology are expanding, transforming how researchers approach the study of ancient humans.
### The Promise of AI in Data Analysis
One of the most significant impacts of AI on paleoanthropology will be in data analysis. AI algorithms can process vast amounts of data far more quickly and accurately than human researchers. This capability is particularly useful in paleoanthropology, where data from fossil records, genetic studies, and archaeological findings can be overwhelming.
#### 1. Automated Fossil Identification
AI-powered tools can automate the identification and classification of fossils. Using machine learning algorithms, these tools can analyze the shape, size, and other characteristics of fossil fragments to determine their species and age. This automation can significantly reduce the time and effort required for fossil analysis, allowing researchers to focus on interpretation and hypothesis testing.
#### 2. Predictive Modeling
AI can also be used to create predictive models of human evolution. By analyzing patterns in fossil records and genetic data, AI algorithms can generate hypotheses about how different hominin species evolved and migrated over time. These models can help researchers identify gaps in the fossil record and guide future excavations.
### Enhanced Imaging and Reconstruction Techniques
The use of AI in imaging and reconstruction techniques is another area where significant advancements are expected. AI can enhance the resolution of imaging technologies, such as CT scans and MRI, and assist in the reconstruction of incomplete fossils.
#### 1. Improved Imaging Resolution
AI algorithms can enhance the resolution of imaging technologies, allowing researchers to examine fossils in greater detail. This improvement can reveal previously unnoticed features and provide new insights into the morphology and biomechanics of ancient hominins.
#### 2. Digital Reconstruction
AI can assist in the digital reconstruction of incomplete fossils. Machine learning algorithms can predict the missing parts of a fossil based on existing data, creating a more complete picture of ancient species. These reconstructions can be used to create 3D models, which can be studied and shared with researchers worldwide.
### Genetic Analysis and Ancient DNA
The analysis of ancient DNA (aDNA) has become a crucial aspect of paleoanthropology, offering direct insights into the genetic makeup of our ancestors. AI is set to revolutionize this area by enhancing the accuracy and efficiency of genetic analysis.
#### 1. AI-Driven Sequencing
AI algorithms can improve the accuracy of DNA sequencing, particularly when working with degraded or contaminated samples. These algorithms can identify and correct errors in sequencing data, providing more reliable results.
#### 2. Comparative Genomics
AI can also be used to compare aDNA with modern human genomes. Machine learning algorithms can identify genetic similarities and differences, shedding light on the evolutionary relationships between ancient and modern humans. This analysis can reveal how certain genetic traits have evolved and spread through populations over time.
### Fieldwork and Excavation
AI is not limited to laboratory settings; it is also making its way into the field. AI-driven technologies can assist in excavation and fieldwork, making these processes more efficient and less invasive.
#### 1. Remote Sensing and Drones
Drones equipped with AI-powered remote sensing technology can survey archaeological sites from the air, identifying areas of interest that may contain fossils or artifacts. This approach can reduce the need for extensive ground surveys and help researchers focus their efforts on promising locations.
#### 2. Robotic Excavation
AI-driven robots can assist in the excavation of delicate fossils. These robots can be programmed to carefully remove soil and debris, reducing the risk of damaging valuable specimens. Additionally, robotic systems can operate in harsh or inaccessible environments, expanding the range of potential excavation sites.
### Educational Outreach and Public Engagement
AI’s impact on paleoanthropology extends beyond research; it also has the potential to transform educational outreach and public engagement. AI-powered tools can make the field more accessible and engaging to a broader audience.
#### 1. Virtual Reality (VR) and Augmented Reality (AR)
AI can enhance VR and AR experiences, allowing users to explore ancient landscapes and interact with digital reconstructions of fossils. These immersive experiences can be used in educational settings to teach students about human evolution and in museums to engage the public.
#### 2. Interactive Platforms
AI can power interactive platforms that allow users to explore paleoanthropological data. These platforms can include features such as virtual excavation sites, interactive timelines of human evolution, and AI-driven chatbots that answer questions about ancient humans. Such tools can make learning about paleoanthropology more interactive and engaging.
### Ethical Considerations and Challenges
While the integration of AI in paleoanthropology holds great promise, it also raises important ethical considerations and challenges. Researchers must navigate these issues to ensure that AI is used responsibly and ethically.
#### 1. Data Privacy
The use of AI in genetic analysis raises concerns about data privacy. Researchers must ensure that genetic data, particularly from indigenous populations, is used with appropriate consent and protection. Ethical guidelines and regulations must be established to govern the use of genetic data in paleoanthropological research.
#### 2. Bias in AI Algorithms
AI algorithms can inherit biases present in the data they are trained on. In paleoanthropology, this could lead to skewed interpretations of human evolution. Researchers must be vigilant in identifying and addressing potential biases in AI algorithms to ensure accurate and unbiased results.
#### 3. Accessibility and Inclusivity
The use of AI in paleoanthropology should be inclusive and accessible to researchers worldwide. Efforts must be made to ensure that AI tools and technologies are available to researchers in developing countries, promoting global collaboration and knowledge sharing.
### Conclusion: A Transformative Decade Ahead
The next ten years promise to be a transformative period for paleoanthropology, driven by the integration of AI technologies. From automated data analysis and enhanced imaging to genetic analysis and robotic excavation, AI is set to revolutionize the field, offering new tools and insights that will deepen our understanding of human evolution.
As researchers navigate the ethical considerations and challenges associated with AI, they must strive to use these technologies responsibly and inclusively. By doing so, they can unlock the full potential of AI to uncover the secrets of our ancient ancestors and share this knowledge with the world.
The future of paleoanthropology is bright, and AI will undoubtedly play a central role in shaping its direction. As we embark on this exciting journey, we can look forward to a decade of groundbreaking discoveries and advancements that will forever change our understanding of human history.
Note – this was written by ChatGPT – 4o
Latest Episode of the PaleoPost Podcast is here!
Rockin’ with Dr. Mazel: Decoding Ancient Art in Africa and Beyond
Please, take a moment to listen, like, and subscribe!
I know you will learn something and enjoy!
Until next time!
WTH
What is the future of Paleoanthropology?
Paleoanthropology, the study of human evolution and our ancient ancestors, continues to be a fascinating field with ongoing discoveries and advancements.
Let’s explore some recent developments and potential future directions:
- Genetic Insights:
- Advances in DNA sequencing technology have allowed researchers to extract and analyze ancient DNA from fossils. For instance, in 2022, a team led by Laurits Skov and Benjamin Peter from the Max Planck Institute for Evolutionary Anthropology sequenced nuclear, mitochondrial, and Y-chromosome DNA from Neanderthal individuals¹[1].
- These genetic studies provide insights into our evolutionary history, relationships between ancient hominins, and even family dynamics among Neanderthals.
- Cutting-Edge Imaging Techniques:
- Modern paleoanthropology benefits from state-of-the-art imaging technology, 3D modeling, and virtual reconstruction. These tools allow scientists to examine fossils in unprecedented detail²[2].
- High-resolution scans help reveal hidden features, such as dental structures or cranial anatomy, shedding light on our ancestors’ lives.
- Diet and Behavior:
- Researchers continue to investigate the diets and behaviors of early humans. Recent discoveries have challenged assumptions about meat consumption and brain evolution.
- For example, a study in 2022 questioned whether increased meat-eating directly correlated with larger brains. It turns out that the relationship is more complex than previously thought¹[1].
- Additionally, evidence of early cooking practices has been pushed back to around 600,000 years ago, suggesting that our ancestors were using fire to cook fish¹[1].
- Environmental Context:
- Paleoanthropologists are increasingly interested in understanding the environments in which our ancient relatives lived. Climate, vegetation, and landscape play crucial roles in shaping human evolution.
- By combining fossil evidence with geological and ecological data, researchers gain a more holistic view of our ancestors’ lives.
- Discovery of New Species:
- The field remains dynamic, with ongoing excavations and fossil discoveries. In 2023, intriguing findings included a 300,000-year-old chinless jawbone from eastern China that displays both modern and archaic features³[6].
- Such discoveries challenge existing species classifications and expand our understanding of human diversity.
- Interdisciplinary Approaches:
- Paleoanthropology increasingly integrates multiple disciplines, including archaeology, geology, paleoecology, comparative anatomy, and experimental biology.
- Collaborative efforts enhance our ability to reconstruct the past and address complex questions about human origins.
- Speculation on Posthumanity:
- While not strictly scientific, speculative works like Arthur C. Clarke’s “2001: A Space Odyssey” explore the prehistory of posthumanity. These imaginative narratives provoke thought about our future evolution⁴[4].
In summary, the future of paleoanthropology lies in continued interdisciplinary research, technological advancements, and the discovery of new fossils. As we uncover more about our ancient past, we gain deeper insights into what it truly means to be human. 🌟¹[1] ²[2] ³[6]
Source: Conversation with Bing, 5/2/2024
(1) Fourteen Discoveries Made About Human Evolution in 2022. https://www.smithsonianmag.com/smithsonian-institution/fourteen-discoveries-made-about-human-evolution-in-2022-180981344/.
(2) How New Tech for Ancient Fossils Could Change The Way We Understand …. https://www.smithsonianmag.com/innovation/new–tech-ancient-fossils-180951647/.
(3) A new human species? Mystery surrounds 300,000-year-old fossil – Nature. https://www.nature.com/articles/d41586-023-02924-8.
(4) Paleoanthropology of the future: the prehistory of posthumanity in …. https://research.monash.edu/en/publications/paleoanthropology-of-the-future-the-prehistory-of-posthumanity-in.
(5) 1 The Past, Present and Future of CHAPTER Paleoanthropology – Wiley. https://catalogimages.wiley.com/images/db/pdf/9781444331165.excerpt.pdf.
(6) Thirteen Discoveries Made About Human Evolution in 2023. https://www.smithsonianmag.com/smithsonian-institution/thirteen-discoveries-made-about-human-evolution-in-2023-180983512/.
Understanding the ‘Muddle in the Middle’ Part 2-Hominins in Asia and Confusion in China-Guest Post by Mekhi

Introduction
During the Pleistocene epoch, the world was full of human species, all throughout Africa, Europe, and Asia, and eventually, North America. Human species were interacting with, living with, and even breeding with each other. Some had vast continent-wide homes, while some migrated far away into new unexplored lands, such as obscure islands in Indonesia or the Philippines. It was a fascinating time, and is one heavily studied by anthropologists today. There is lots of fossil and genetic evidence providing clear pictures of most of these species, but there are also many gaps in our knowledge.
The many fossils we have are very useful for understanding this time in human history, but also can cause lots of confusion, mainly in terms of taxonomy. In a previous article (https://worldofpaleoanthropology.org/2023/08/23/understanding-the-muddle-in-the-middle-hominins-from-the-pleistocene-guest-post-by-mekhi/), I discussed the Pleistocene hominins from Europe and some from Africa, and the confusion that comes with them, but recent research has added more confusion to the other species, this time living farther east, in Asia.
There were many hominin species in Asia, and they arguably cause more confusion than what is seen in Africa and Europe. To understand this, we first need to go through all the known human species from the area.
Hominin Diversity and Distribution During the Pleistocene of Asia
Animals often have large geographical ranges, and live in more than one place. Leopards (Panthera pardus) for example, have a huge range, mostly living in Africa, but also living in Europe and Asia, having the biggest range of any big cat today. Some species of humans had similarly large ranges. Obviously, Homo sapiens today have a massive range, living pretty much across the whole globe, but even other extinct species had large ranges as well. It is important to note a big difference between humans and leopards in this comparison though, which is that though leopards have a broad geographic range, the species is split into different subspecies in different areas, whereas other extinct human species are typically not.
A good example of broad geographic ranges is Homo heidelbergensis from Africa and Europe, though there has been contention on whether the African populations were the same species or not. Another species, one more relevant to this topic, is Homo neanderthalensis, or the Neanderthals, known from both Europe and (sparsely) from Asia.
Neanderthals in Asia
Neanderthals were mostly settled in Europe, but were also present in Asia, mainly western Asia and the middle east, but sites containing their remains are also known farther east from Russia. Because it can sometimes be tricky to identify whether a specimen actually belonged to a Neanderthal or not, especially one so far from home, genetic analysis is often used. Genetic research on hominin remains in Russia shows that Neanderthals were present as far as the Altai region of Siberia.
A cave called Chagyrskaya in this region is very famous for Neanderthals. This site, dated to ~60,000 years ago (kya), is not only a great example of the Neanderthal remains in Asia, with many tools, Neanderthal bones, and bones of other animals are known from it, but it is also a prime spot for Neanderthal genetics.
Genetic research has shown that this population was relatively isolated, and had less than 60 individuals present, which is still large for Neanderthal standards. The genetic material from this cave is also exceptionally well preserved. The genome of one individual was sequenced for example, showing that she was female and closely related to the other known Neanderthals from western Asia.
Another cave, very close to Chagyrskaya, also documents Neanderthals in Russia; it is known as Okladnikov cave. Nearby to both these caves, housing a very unknown species only found in Asia, is Denisova cave, known for the Denisovans.

Denisovans
The Denisovans were a species of late genus Homo, very similar to Neanderthals, that lived in east Asia ~500-40 kya. Though our knowledge about this species is still growing, they’re still a cause of great confusion, mostly due to the fact that we have very, very few fossils. We have no skeletons, no skulls, nothing, just a few teeth and bone fragments. Almost everything we know about them is known just from genetic material. What we actually can learn about Denisovan morphology comes mostly from the dental remains.
The morphology seen in Denisovan teeth is mostly unique from that of Neanderthals and Homo sapiens, showing their designation as a separate species. The teeth do show similarities with Asian Homo erectus, but the overall morphology is mostly unique.
Though the Denisovans were most likely a distinct species, no true species name has been given due to the lack of fossil material. Species names like Homo altaiensis or Homo denisova may be used in the future.
The Asian Neanderthals and Denisovans also inhabited the same region, at around the same time. Neanderthals and Denisovans were sister species, and shared a recent common ancestor that was itself a sister species to Homo sapiens. Similar to how people can have some levels of Neanderthal DNA within their genomes, people today can also have some Denisovan DNA, with present-day Melenesians having up to 4-6% Denisovan DNAin their genomes. Much of the genetic material we do have of Denisovans to compare this comes from the few physical remains we actually have from them.
Denisovan genetic material is known from multiple remains. The most important and famous example of this may be the individual known as “Denny”. This individual, known from the fragmentary ‘Denisova 11’ remains, has very well preserved genetic material. From these remains, we know the sex of this individual, which is female, her eye color, which is brown, and most importantly, who her parents were. Her father was a Denisovan, but her mother was a Neanderthal, making her a hybrid. She also was ~13 years of age.

This shows that not only were Neanderthals living with and interacting with Denisovans, but they were also breeding with them. This suggests that Neanderthals were in this region by 120 kya, even as old as potentially 190 kya. Very little is known about Denisovan biology, but more is known about their culture.
Evidence of fire use, mainly charcoal and ashes in the sediments, has been found in Denisova cave, suggesting these people were using fire, which isn’t very surprising. Stone tools have also been uncovered in the cave, giving insight to their technology. Multiple stages of occupation can be found from these tools, each from the different and distinct sedimentary layers containing different tool assemblages from different times. Layers 11.3, 13, 14, and 15 come from the middle paleolithic, and layers 11.1 and 11.2 from the upper paleolithic.
Also from the upper paleolithic layers, specifically the initial upper paleolithic, are lots of bone tools and decorative ornaments. These bone tools consist of an eyed needle and awe from the bones of large mammals. The bone ornaments, or jewelry, include beads, pendants, plaques, rings, a button/fastener, and decorated bones and bracelets.
These bone tools show that the Denisovans were smart, and could take advantage of multiple resources. It also shows that they were culturally advanced, and could make jewelry, though simple, to decorate their bodies. It has long been thought jewelry was something unique to modern Homo sapiens, but we now have evidence that other human species including both Denisovans and Neanderthals were doing it too.

The Denisovans were and still are an odd and confusing species, but more species were causing more confusion down in southeast Asia at this time. One such species was Homo floresiensis, also known as the ‘hobbit’ species.
Homo floresiensis
Homo floresiensis at first sounds like a normal hominin species, living on the island of Flores in Indonesia from ~190-50 kya. This species was one of the Pleistocene hominins in Asia, along with Neanderthals, Denisovans, and all the other species that will be discussed later, but there is something unique, and very confusing about this species.
Though very human-like, clearly in the genus Homo, this species was very primitive, especially in the fact that the adults were only ~1 meter (~3 feet) tall. This short stature of this strange species has caused great confusion, and a few different hypotheses have been proposed to explain it.
At first, the fossils, found in Liang Bua cave, were interpreted as pathological modern humans, humans who suffered from microcephaly (a decrease in the size of the head). However, this hypothesis was very quickly shown to be false. The skull of H. floresiensis and that of a microcephalic human are very different, and H. floresiensis lacks the expected features for a person with this condition. This gave a need for new hypotheses. H. floresiensis clearly wasn’t a modern human, nor was it ancestral to modern humans; it was a strange ‘hobbit’ species living on a secluded island in Indonesia.
This idea of an off-branching species was very important at the time; it was commonly thought that human evolution was a straight line of progression, with the only branching members of the lineage being groups like the robust australopiths (Paranthropus) and Neanderthals. This species showed the true diversity of hominins that lived before modern humans that weren’t actually ancestral to modern humans.
The next hypothesis for the origins of these hobbits was that they were a subset of Homo erectus, a species that inhabited the nearby island of Java at the time, that simply suffered from insular dwarfism, a common evolutionary phenomenon that occurs when species are trapped on isolated islands with limited resources. This would explain why H. floresiensis was so short and why it was there, but it would not explain the rest of the morphology.
Even early on in the research of H. floresiensis, scientists noticed that the species had many basal, or primitive characteristics, traits shared with earlier African hominins such as Australopithecus and early Homo. Some of the obvious primitive traits include its short stature, but it also had a small brain volume (~380 cubic centimeters), and limb proportions similar to that of earlier hominins. However, it still had some derived features shared with later species like Homo erectus, such as a flatter, more orthognathic face, and more derived teeth.
Cladistic analysis suggests that H. floresiensis arose sometime around the time of Homo habilis and Homo rudolfensis, the two earliest known members of the genus Homo, though it is possible that it came sometime after early Homo. The origin of H. floresiensis is still highly debated; both ideas are very plausible and have support, but the main consensus right now seems to be that this species is a descendant of early hominin species from Africa, and just happened to end up nearby to where H. erectus would inhabit.
This however does bring up another question, the question of how this species even got to the island in the first place. Flores is an island, and has not recently been in contact with other land masses, unlike other Indonesian islands like Java. This means that H. floresiensis had to cross some waterways to get there. This isn’t actually too implausible. Much of the native wildlife of the island, from mammals to reptiles, seem to have come to the island from neighboring islands, such as Java, Sahul, and Sulawesi. Some of the larger animals, such as large reptiles and the extinct elephant relative Stegodon likely could have swam or even floated, while the smaller mammals, such as rodents, may have rafted there. These are all also species that H. floresiensis ate and butchered.
This rafting hypothesis easily applies to H. floresiensis. We can never know the true path the hobbits took to get there, but they very well may have rafted, likely unintentionally, to the island they would call home. They would have a long time of success on the island after their arrival, but went extinct at some point, and their extinction offers even further confusion.
Natural disasters, such as tsunamis and volcanic events are common there, and definitely had effects on them, but they don’t seem to have wiped them out. A large volcanic eruption is known to have occurred on Flores ~12 kya, but this was too late in time, as H. floresiensis seems to have been extinct by this point. Their disappearance seems to have occurred ~50 kya, which matches exactly with the known arrival date of Homo sapiens on the island.
For a brief period, our species seems to have coexisted with the hobbits, and unfortunately, we may have led to their extinction. The origins and disappearance of these hobbits adds more confusion to what already was a confusing time and place, but they weren’t the only confusing species in the area.

Homo luzonensis
While Homo floresiensis was living in Indonesia, another confusing species was living nearby in the Philippines, called Homo luzonensis. Similar to the Denisovans, H. luzonensis is known from very little fossil material, though we have more from this species than we have from the Denisovans. Unlike the Denisovans however, we have no genetic material from H. luzonensis. No full skeletons have been found either, unlike H. floresiensis. Only 13 remains dated to 67 kya have been uncovered. Overall, very little is known about this species.
The remains from this species include mostly teeth, some finger and toe bones, and the femur of a young child. All the remains come from the same stratigraphic layer of Callao Cave, in the island of Luzon. Though very small in numbers, the fossils we do have can tell us some things, or at least give us hints. The teeth may be the most useful of the remains for learning about this species.
Out of the 13 remains, 7 of them were teeth, 5 of which seem to have belonged to the same individual. These 5 teeth make up the holotype of the species, another reason why they have a species name and the Denisovans don’t. The teeth are primitive and small, and similar to those of H. floresiensis.
The small size of the teeth suggests that H. luzonensis may have had a small body size, as tooth size often reflects body size, though this isn’t always the case. The metatarsals and metacarpals in the feet and hands are also very small and primitive, resembling H. floresiensis and early members of the genus like Homo habilis. Despite the size of the teeth, they also share many traits with Homo erectus. This could suggest that H. luzonensis descended from H. erectus and potentially suffered some insular dwarfism, though it is possible that it was a descendant of earlier hominins, like what is thought for H. floresiensis.
Like H. floresiensis, the origins of H. luzonensis are unknown and heavily debated, though, also like H. floresiensis, whoever they descended from, they likely got to the island via rafting, as Luzon has never been recently in contact with any other landform. If not from earlier hominin species, these two island based human species must have descended from H. erectus, the only other hominin species living in the area.
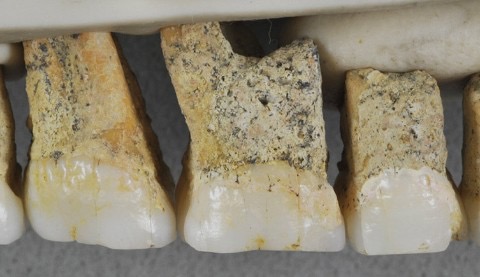
Homo erectus
Homo erectus was perhaps the most successful human species, at least for its time. The oldest H. erectus fossils are known from 1.8 million years ago (mya), to as recent as ~100 kya. H. erectus arose out of Africa, and migrated all throughout Eurasia, settling mostly in southeast Asia.
The site known as Sangiran, in the Solo basin on the island of Java Indonesia has revealed over 40 H. erectus specimens, making it perhaps the most important locality for H. erectus in Indonesia. Sangiran is one of the 3 main sites known for Homo erectus remains in Eurasia, with the others being Dmanisi in eastern Europe and Zhoukoudian in China. The earliest of these fossils in Indonesia are about 1.5m years old, which is very close to the 1.8m years old remains in Dmanisi, the oldest H. erectus remains out of Africa. This shows that they spread and migrated very quickly.
This population in Indonesia however lived for a very long time after that; they’re seemingly the last surviving group of H. erectus. 12 H. erectus skull caps, along with 2 leg bones, uncovered from Ngandong, central Java, near the Solo River, represent the last known H. erectus population to have lived, along with the most derived population. These fossils lived between 117-108 kya, showing truly just how long H. erectus lived as a species.
This brings up the question of how did H. erectus reach Java in the first place? The answer to that is glaciation. During the Pleistocene, as old as 2.6 mya, glaciations resulted in drastic drops in sea level, as much as 72-120 meters (~236-393 feet) below current sea level in some cases. This resulted in many land bridges across Indonesia, allowing for faunal migration to the islands. This led to the colonization of Java by many different animals by about 2 mya, including mammoths and likely H. erectus, though it is likely that they still had to cross some water bodies to get there.
When they arrived at the island, they were introduced to an environment of moist grasslands and open woodlands, with riverine landscapes and large streams, along with long annual dry seasons. Aside from Indonesia, H. erectus is also commonly known from China, mainly at a site called Zhoukoudian.
Zhoukoudian represents ~80 individuals, the most number of H. erectus individuals known from anywhere, along with ~100,000 stone tools. This population arrived there long after others arrived in Indonesia, living there from ~750-200 mya, though their occupation there seems to have been sporadic.
Unfortunately, by 108k years, the all-mighty Homo erectus had gone extinct. It is hypothesized that their extinction was caused by climate and environmental changes in the region. The vast and open environments they migrated into, found in both Indonesia and China, transitioned to a wet and crowded rainforest, which may have ended their reign. However, we likely will never know what truly wiped them out, as it is hard to point at the exact causes of small extinctions.
H. erectus is the cause of little confusion, being one of the better understood species from the Pleistocene. However, there was another human population living in China that is incredibly unknown, unique and confusing, more so than any of the other species of the time, making the name the ‘muddle in the middle’ especially well deserved.

Homo longi
The next species in Asia is called Homo longi. This species is the most recently named human species, being first described in 2021. Even though it was first described in 2021, the skull, known as the Harbin skull, was originally discovered in 1933 during a construction project in the Hebei Province of northern China, although the exact locality is unknown. The skull was dated to ~146 kya.
The skull itself is one of the most well preserved hominin crania from the Pleistocene of Asia; it is only missing most of its teeth and possesses damage to its left zygomatic arch. It is also very large. It has a large cranial capacity of ~1,420 cubic centimeters. Some of its morphology is similar to that of Homo sapiens, such as its large cranial capacity, short face, and small cheekbones, but it also possesses some other unique morphology. This includes a low cranial vault, pronounced brow ridges, large molars, and alveolar prognathism below the nose.
It also shares features with Homo heidelbergensis and Neanderthals. This is a unique and strange combination of traits. This set of traits is also seen in other hominin crania from China, that seem to represent a unique and unknown human population or species living in China during this time, that includes Homo longi. As it turns out, there is a lot of hominin fossil material from the Pleistocene from all throughout China, and these fossils are the most confusing part of human evolution in the Pleistocene of Asia.

The Mysterious Hominins of Pleistocene China
Much of the hominin fossil record of Pleistocene China, just as it is as a whole, is teeth. Below is a chart showing different locations in China where hominin teeth were uncovered, spanning a temporal range of 990-15 kya.
| Location of Discovery | Dating | Number of Teeth |
| Meipu, southern China | 990-780 KYA | 4 |
| Yiyuan, eastern China | 770-126 KYA | 7 |
| Longtan, Cave, Hexian, eastern China | 412 KYA | 10 |
| Panxian Dadong, southern China | 300-130 KYA | 4 |
| Tongzi, southern China | 172-240 KYA | 4 |
| Luna Cave, southern China | 126-70 KYA | 2 |
| Dushan Cave, southern China | 15 KYA | 1 |
Only a few of the dental remains have been assigned a species, such as the teeth from Luna cave, which clearly belong to Homo sapiens. The morphology of these teeth is also interesting. Many of the teeth have Homo erectus-like features, while other traits appear more modern. This mosaic morphology in the dentition is similar to the mosaic morphology of the rest of the bodies, especially the skulls.
This mosaic nature, the mix of archaic and modern features has caused some to suggest that many of these remains belong to hybrid individuals, hybrids of archaic and modern human species, such as H. erectus and H. sapiens. This idea has especially been proposed for some of the postcranial material that has been uncovered, mainly femora.
The skulls from this time and place, as mentioned, also display strange morphology, and have been under great research lately, revealing a lot about the hominins living in China. A collection of well preserved hominin crania, including the Harbin skull of Homo longi, make up the best set of hominin remains from this time, and are the most useful and important. Well preserved hominin crania are known from 5 sites, Harbin, Dali, Jinnisuan, Maba, and maybe most importantly, Hualongdon.
Dali
The Dali skull, from the Shaanxi Province of northwest China, was originally uncovered in 1978. This skull has been estimated to be ~200k years old. The specimen is mostly complete, only having some distortion and a few missing pieces, including a big chunk of the parietal bone.
The face of the Dali skull resembles that of Homo sapiens, but aside from the face, the skull resembles other African and eastern Eurasian Pleistocene hominins. Overall, it most greatly resembles the skulls of early H. sapiens of north Africa and the Levant region. This combination of traits from multiple hominins is a consistent trend throughout all these skulls.
The Dali skull may show that during the Pleistocene, many different human species and populations in Asia were breeding with one another, creating a ‘braided stream’, with gene flow from different groups creating unique morphology in different populations. At its extremes, this idea is known as the ‘multiregional hypothesis’, and is in contention with the theory of ‘out of Africa’.


Jinniushan
The next important specimen is not not just a skull, but a whole partial skeleton and cranium from northeast China. This is very rare in the hominin fossil record from this time, as most specimens are either just a skull, or fragmentary remains like teeth or small postcranial (body) material. This is not the case for the Jinniushan specimen, which consists of a skull, along with an arm bone (left ulna), a pelvic bone (left innominate), 6 vertebrae, ribs, and bones from the hands and the feet.
The skull itself is fairly complete, but was broken during its excavation in 1984. What is missing from the skull is mostly from the frontal, parietal, and occipital regions. The dating of this specimen has been somewhat controversial, but current estimates place it at roughly ~200 kya, similar in time to Dali. Other fossils come from the same strata as the Jinniushan specimen, including bears, wolves, deer, and rhinos.
Morphologically, the skull shares some similarities with Dali, along with a mix of Homo erectus and Homo sapiens features. They both possess a small mastoid process, similar parietal bones, and a shared location of the maximum cranial breadth. There are many differences too, including smaller brow ridges, a shallower supraorbital sulcus, etc. The cranial capacity of the Jinnushan skull is roughly 1,400 cubic centimeters, which is very large, and around the same size as modern H. sapiens.
The presence of well preserved postcranial material provides a rare case in which the encephalization quotient (the size of the brain compared to the body) can be seen. In Jinniushan, the encephalization quotient is 4.15, which is very close to the predicted encephalization quotient for other pleistocene hominins from this area. As for the rest of the body, this individual’s body mass was estimated to be 78.6 kg (~172 lbs), and the body height to be 168 cm (~5 ½ ft). It also had shorter limbs than expected.
All these body proportions are consistent with adaptations for colder weather, in which species adapt to be larger and have shorter limbs to conserve body warmth. This is known as Allen’s rule, and is also seen in other hominin species such as Neanderthals. The climate in China during the Pleistocene is not extensively studied, but research has revealed a climate transition, known as the Mid-Pleistocene climate transition (MPT), characterized by increased monsoon and glaciation cycles from ~1.2m to 700 kya. This occurred before the Jinniushan specimen was around, but the presence of these features may suggest similar conditions during its time.
The foot of the Jinniushan specimen is also interesting. It possessed some derived features, such as an increased stability of the medial longitudinal arch, but it also had several primitive features, such as lower arches and a less stable hallucal metatarsophalangeal joint. These traits provide a glimpse into the stride of this pleistocene hominin; they suggest that this individual had a slightly different stride than what is seen in modern H. sapiens.
Other details about the Jinniushan specimen include that it was likely female based off of the pelvis, and was ~16-20 years of age, making her relatively young. Stone tools and traces of fire have also been reported alongside the Jinniushan specimen.
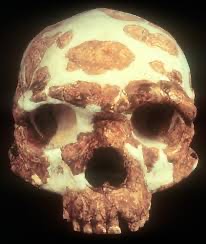
Maba
After Jinniushan in this skull set comes the Maba skull. This skull, from ~300-130 kya is very fragmentary, consisting of a skull cap, a right orbit, and a portion of the nasal bone. Along with this, it possesses pronounced brow ridges and a heavily curved frontal bone, more so than Chinese Homo erectus. The brow ridges are also similar in shape and size to European Neanderthals.
The thickness of the cranial vault is also similar to that of modern Asian Homo sapiens. Just as with all the other crania discussed so far, Maba possesses a strange mosaic of features shared between multiple other hominin species. It is also somewhat unique from Dali and Jinniushan, having a more prognathic face. The cranial capacity of Maba is about 1,300 cubic centimeters, within the ranges of Homo heidelbergensis, Neanderthals, and H. sapiens. There are a few other hominin remains aside from the cranium as well, consisting of 5 teeth and a partial mandible, though they seem to not be associated with the Maba cranium, as they dated to ~237 kya, and some of the teeth are associated with H. sapiens instead.


The last strange skull that will be discussed is one of the more recently described specimens, like Harbin. It may be one of the more important specimens known from this time and place; it is the HLD6 Hualongdong skull.
Hualongdong
The taxonomic placement of all these skulls is very confusing. They’re all unique in their own ways, and all have mosaic characteristics, combining multiple traits from several other hominins. One commonly thought idea is that these skulls, or at least for some of them, are Denisovans. They’re in the right place and time, but this could never be confirmed.
We have no confirmed Denisovan cranial material; there is nothing to definitively back up the claim that these Chinese skulls represent Denisovans, and no genetic material from these specimens to compare. There is the Denisovan partial mandible, Xiahe, but no cranium, and all of the skulls discussed here are crania, but have no associated mandibles, at least up until Hualongdong.
The specimen known as HLD 6, from Hualongdong eastern China, is a ~300k year old juvenile hominin skull and mandible, along with some dental remains. The skull’s face resembles that of other Asian hominin crania, including Dali, Jinniushan, Maba, and even Homo erectus. The face of the skull is very flat, like Homo sapiens, though this could also be because the individual was young, only ~13-15 years of age; face shape changes lots in hominin ontogeny. Overall, the cranium fits in well with the other mosaic skulls from this time and place. However, the mandible is the more important part of the specimen.

The mandible is broadly characterized by a robust mandibular corpus (the body of the mandible), and a more gracile mandibular symphysis and ramus (the front and back ends of the mandible). It also lacks a chin, showing that it can’t belong to H. sapiens. It does however share many other traits with other pleistocene hominin mandibles, including a pronounced alveolar planum, a superior transverse torus, a pronounced endocondyloid crest, and a well-developed medial pterygoid tubercle.
It shares lots in common with other Pleistocene hominins from throughout Eurasia. This is significant as it is one of the only complete hominin mandibles from China, only going alongside Penghu 1 from Taiwan (~190-10 kya), Xujiayao 14 from northern China (~120 kya), and the Denisovan Xiahe mandible (~160 kya).
Despite also being from China, the HLD6 mandible differs greatly from these other mandibles. This is significant as no other of the Chinese crania discussed so far also have associated mandibles. Because the HLD6 cranium is so similar to the other crania, it is reasonable to assume that its mandible is similar as well. If the mandibles are similar, it means that all the mandibles are distinct from the other Chinese mandibles.
This is especially important for the Xiahe mandible. If the Xiahe mandible is distinct from the other mandibles, then the cranium may be too. This could imply that the Denisovan cranium and these other Chinese crania are distinct, showing that these hominins may not be Denisovans. This especially applies to Homo longi, as it has been long speculated to be Denisovan. This could be a huge discovery, but it is very important to note that this claim is entirely based on logical speculation, and will need significant further, physical evidence to be proven.

What Are These Skulls?
So now we know what the skulls (probably) aren’t, what exactly are they? These fossils are clearly not modern Homo sapiens, and also shouldn’t be grouped in as Homo erectus either, despite its presence in the region at this time. One species that these hominins have sometimes been grouped under is Asian Homo heidelbergensis.
Homo heidelbergensis already has had a history of confusion when it comes to many specimens being grouped into it that may not necessarily belong there. Though many of the African and European hominin specimens fit into H. heidelbergensis, the Asian specimens seem to be overall distinct.
The 2 main reasons why these Asian specimens are sometimes grouped into H. heidelbergensis are 1) some apomorphic (derived) traits seen in Asia are also found in European H. heidelbergensis, and 2) Homo sapiens didn’t descend from Asian Homo erectus, so it must be H. heidelbergensis instead.
Some of the shared traits include very heavily increased encephalization, and the presence of canine fossa, which is found in several of the Chinese specimens and also the fossils from Gran Dolina Spain, associated with Homo antecessor. This shows that the Chinese specimens may represent a group closer to H. heidelbergensis cladistically, and farther from H. erectus.
However, despite these shared traits, there are many distinguishing traits as well. These traits include the orientation of the frontosphenoidal process of the zygomatic bone, the upper facial height, shovel shaped incisors, the presence of interparietal bones, and agenesis of the 3rd molars. Some H. heidelbergensis do possess some of these traits, such as interparietal bones, but they’re uncommon.
Overall, it is unlikely that the Chinese specimens represent H. heidelbergensis, but they may be related to it. Another common idea for the classification of these specimens is placing them in a new species. Homo longi is the only one of the Chinese crania that has an actual assigned species, but other species names have been proposed for other specimens, such as Homo daliensis and Homo mabaensis. This however, is counterproductive; the Pleistocene is already very confusing in terms of hominin taxonomy with lots of very confusing taxa; adding more taxa will only muddle it up more.
It might not be terrible to group them all under one diverse species, such as M. mabaensis, but all this would do would be changing what we call the fossils, and wouldn’t do much for our actual understanding. It has been proposed that the safest course of action in terms of what to call these specimens is simply to refer to them as archaic Homo sapiens until further analysis clears up the muddle in Asia.
Conclusion
This mysterious group of hominins clearly were a distinct group, likely closely related to H. heidelbergensis and other Pleistocene hominins. Classifying these hominins is difficult, as there isn’t any consistent set of traits that define them. The only truly defining trait is the increasing morphological diversity throughout all of them. It is also unknown how these hominins ended up where they did. There are 3 hypotheses for how they got to China, but nothing for certain.
The first idea is that they got there via the Chao Phraya river basin; the second is that they got there through east Vietnam, and the 3rd and last idea is that they traveled along the coast of southern Asia and Myanmar. These are all plausible ideas, but none of them have enough evidence to back them up to make them certain.
Until more evidence and research comes along, the Pleistocene of Asia will remain confusing, and, just like with Europe, further research and discoveries, such as new fossils and genetic evidence are our best hope to resolve this confusing time.
Sources
- “Where Do Leopards Live? In the Wild, Of Course – Right?” africafreak (ND). https://africafreak.com/where-do-leopards-live
- Dorey, Fran. “Homo heidelbergensis”. The Australian Museum, 06-28-21.https://australian.museum/learn/science/human-evolution/homo-heidelbergensis/
- Dorey, Fran. “Homo neanderthalensis”. The Australian Museum, 06-28-21. https://australian.museum/learn/science/human-evolution/homo-neanderthalensis/
- Krause, J., Orlando, L., Serre, D., Viola, B., Prüfer, K., Richards, P. M., Hublin, J., Hänni, C., Derevianko, P. A., Pääbo, S. (2007). Neanderthals in central Asia and Siberia. Nature, 499, 902-904. https://doi.org/10.1038/nature06193
- Salazar-García, C. D., Power, C. R., Rudaya, N., Kolobova, K., Markin, S., Krivoshapkin, A., Henry, G. A., Richards, P. M., Viola, B. (2021). Dietary evidence from Central Asian Neanderthals: A combined isotope and plant microremains approach at Chagyrskaya Cave (Altai, Russia). Journal of Human Evolution, 156, 102589. https://doi.org/10.1016/j.jhevol.2021.102985
- Mafessoni, F., Grote, S., de Filippo, C., Pääbo, S. (2020). A high-coverage Neandertal genome from Chagyrskaya Cave. PNAS, 11(26): 15132-15136. https://doi.org/10.1073/pnas.2004944117
- Dobrovolskaya, V. M., Tuinov, V. A. (2013). The Neanderthals of Okladnikov Cave Altai: Environment and Diet Based on Isotopic Analysis. Archaeology, Ethnology, and Anthropology of Eurasia. 41(1): 78-88. https://doi.org/10.1016/j.aeae.2013.07.007
- Dorey, Fran. “The Denisovans”. The Australian Museum, 20-04-20. https://australian.museum/learn/science/human-evolution/the-denisovans/
- Zubova A. V., Chikisheva T .A., Shunkov M. V. (2017). The Morphology of Permanent Molars from the Paleolithic Layers of Denisova Cave. Archaeology, Ethnology & Anthropology of Eurasia. 45(1): 121-134. https://doi.org/10.17746/1563-0110.2017.45.1.121-134
- Jacobs Z., Li, B., Shunkov, V. M., Kozlikin, B. M., Bolikovskaya, S. N., Agadjanian, K. A., Uliyanov, A. V., Vasiliev, K. S., O’Gorman, K., Derevianko, P. A., Roberts, G. R. (2019). Timing of archaic hominin occupation of Denisova Cave in southern Siberia. Nature, 565, 594-599. https://doi.org/10.1038/s41586-018-0843-2
- Reich, D., Green, E. R., Kircher, M., Krause, J., Patterson, N., Durand, Y. E., Viola, B., Briggs, W. A., Stenzel, U., Johnson, F. L. P., Maricic, T., Good, M. J., Marques-Bonet, T., Alkan, C., Fu, Q., Mallick, S., Li, H., Meyer, M., Eichler, E. E., Stoneking, M., Richards, M., Talamo, S., Shunkov, V. M., Derevianko, P. A., Pääbo, S. (2010). Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature, 468, 1053-1060. https://doi.org/10.1038/nature09710
- Slon, V., Mafessoni, F., Vernot, B., de Filippo, C., Grote, S., Viola, B., Hajdinjak, M., Peyrégne, S., Nagel, S., Brown, S., Douka, K., Higham, T., Kozlikin, B. M., Shunkov, V. M., Derevianko, P. A., Kelso, J., Meyer, M., Prüfer, K., Pääbo, S. (2018). The genome of the offspring of a Neanderthal mother and a Denisovan father. Nature, 561, 113-116. https://doi.org/10.1038/s41586-018-0455-x
- Morely, W. M., Goldberg, P., Uliyanov, A. V., Kozlikin, B. M., Shunkov, V. M., Derevianko, P. A., Jacobs, Z., Roberts, G. R. (2019). Hominin and animal activities in the microstratigraphic record from Denisova Cave (Altai Mountains, Russia). Scientific Reports, 9, 13785. https://doi.org/10.1038/s41598-019-49930-3
- Shunkov, V. M., Kozlikin, B. M., Derevianko, P. A. (2020). Dynamics of the Altai Paleolithic industries in the archaeological record of Denisova Cave. Quaternary International, 559, 34-46. https://doi.org/10.1016/j.quaint.2020.02.017
- Shunkov, V. M., Fedorchenko, U. A., Kozlikin, B. M., Derevianko, P. A. (2020). Initial Upper Palaeolithic ornaments and formal bone tools from the East Chamber of Denisova Cave in the Russian Altai. Quaternary International, 559, 47-67. https://doi.org/10.1016/j.quaint.2020.07.027
- O’Donoghue, Saskia. “Ancient human DNA found in 25,000-year-old tooth jewelry”. Euronews, 08-05-2023. https://www.euronews.com/culture/2023/05/08/ancient-human-dna-found-in-25000-year-old-tooth-jewellery
- Dorey, Fran. “Homo floresiensis”. The Australian Museum, 23-12-19. https://australian.museum/learn/science/human-evolution/homo-floresiensis/
- Groves, C. (2007). The Homo floresiensis Controversy. Hayati Journal of Biosciences, 14(4): 123-126. https://doi.org/10.4308/hjb.14.4.123
- Argue, D., Morwood, J. M., Sutkina, T., Jatmiko, Saptomo, W. E. (2009). Homo floresiensis: a cladistic analysis. Journal of Human Evolution, 57(5): 623-639. https://doi.org/10.1016/j.jhevol.2009.05.002
- Aiello, C. L. (2010). Five years of Homo floresiensis. American Journal of Biological Anthropology, 142(2): 167-179. https://doi.org/10.1002/ajpa.21255
- Brumm, A., Aziz, F., van den Bergh, D. G., Morwood, J. M., Moore, W. M., Kurniawan, I., Hobbs, R. D., Fullagar, R. (2006). Early stone technology on Flores and its implications for Homo floresiensis. Nature, 441, 624-628. https://doi.org/10.1038/nature04618
- Dennell, W. R., Louys, J., O’Reagan, J. H., Wilkinson, M. D. (2014). The origins and persistence of Homo floresiensis on Flores: biogeographical and ecological perspectives. Quaternary Science Reviews, 96, 98-107. https://doi.org/10.1016/j.quascirev.2013.06.031
- Davis, Josh. “Homo luzonensis: new species of ancient human discovered in the Philippines”. Natural History Museum, 04-11-19. https://www.nhm.ac.uk/discover/news/2019/april/new-species-of-ancient-human-discovered-in-the-philippines.html
- Détroit, F., Mijares, S. A., Corny, J., Daver, G., Zanolli, C., Dizon, E., Robles, E., Grün, R., Piper, J. P. (2019). A new species of Homo from the Late Pleistocene of the Philippines. Nature, 568, 181-186. https://doi.org/10.1038/s41586-019-1067-9
- Zanolli, C., Kaifu, Y., Pan, L., Xing, S., Mijares, S. A., Kullmer, O., Schrenk, F., Corny, J., Dizon, E., Robles, E., Détroit, F. (2022). Further analyses of the structural organization of Homo luzonensis teeth: Evolutionary implications. Journal of Evolution, 163, 103124. https://doi.org/10.1016/j.jhevol.2021.103124
- Roberts, L. D., Jarić, I., Lycett, J. S., Flicker, D., Key, A. (2023). Homo floresiensis and Homo luzonensis are not temporally exceptional relative to Homo erectus. Journal of Quaternary Science, 38(4): 463-470. https://doi.org/10.1002/jqs.3498
- Dorey, Fran. “Homo erectus”. The Australian Museum, 16-10-20. https://australian.museum/learn/science/human-evolution/homo-erectus/
- Zaim, Y., Ciochon, L. R., Polanski, M. J, Grine, E. F., Bettis, III, A. E., Rizal, Y., Franciscus, G. R., Larick, R. R., Aswan, H. M., Eaves, L. K., Marsh, E. H. (2011). New 1.5 million-year-old Homo erectus maxilla from Sangiran (Central Java, Indonesia). Journal of Human Evolution, 61(4): 364-376. https://doi.org/10.1016/j.jhevol.2011.04.009
- Widianto, H., Noerwidi, S., Hascaryo, T. A. (2023). New Hominin calvaria discovery from Grenzbank Layer of Sangiran Dome (Java, Indonesia): The last archaic Homo erectus lived in Java. L’Anthropologie, 127(3): 103165. https://doi.org/10.1016/j.anthro.2023.103165
- Rizal, Y., Westaway, E. K., Zaim, Y., van den Bergh, D. G., Bettis, III, A. E., et al. (2019). Last appearance of Homo erectus at Ngandong, Java, 117,000–108,000 years ago. Nature, 577, 381-385. https://doi.org/10.1038/s41586-019-1863-2
- Widianto, H., Noerwidi, S. (2023). Long journey of Indonesian Homo erectus: Arrival and dispersal in Java Island. L’Anthropologie, 127(3): 103167. https://doi.org/10.1016/j.anthro.2023.103167
- Bettis, III, A. E., Milius, K. A., Carpenter, J. S., Larick, R., Zaim, Y., Rizal, Y., Ciochon, L. R., Tassier-Surine, A. S., Suminto, M. D., Bronto, S. (2009). Way out of Africa: Early Pleistocene paleoenvironments inhabited by Homo erectus in Sangiran, Java. Journal of Human Evolution, 56(1): 11-24. https://doi.org/10.1016/j.jhevol.2008.09.003
- Choi, Q. Charles. “The Final Days of Homo Erectus”. Science Insider.12-18-19. https://www.insidescience.org/news/final-days-homo-erectus#
- Ni, X., Ji, Q., Wu, W., Li, Q., Grün, R., Stringer, C. (2021). Massive cranium from Harbin in northeastern China establishes a new Middle Pleistocene human lineage. The Innovation, https://doi.org/10.1016/j.xinn.2021.100130
- Liu, W., Schepartz, A. L., Xing, S., Miller-Antonio, S., Wu, X., Trinkaus, E., Martinón-Torres, M. (2013). Late Middle Pleistocene hominin teeth from Panxian Dadong, South China. Journal of Human Evolution, 64(5): 337-355. https://doi.org/10.1016/j.jhevol.2012.10.012
- Xing, S., Martinón-Torres, M., Bermúdez de Castro, M. J., Zhang, Y., Fan, X., Zheng, L., Huang, W., Liu, W. (2014). Middle Pleistocene Hominin Teeth from Longtan Cave, Hexian, China. PLoS ONE, https://doi.org/10.1371/journal.pone.0114265
- Xing, S., Martinón-Torres, M., Deng, C., Shao, Q., Wang, Y., Luo, Y., Zhou, X., Pan, L., Ge, J., Bermúdez de Castro, M. J., Liu, W. (2021). Early Pleistocene hominin teeth from Meipu, southern China. Journal of Human Evolution, 151, 102924. https://doi.org/10.1016/j.jhevol.2020.102924
- Xing, S., Martinón-Torres, M., Bermúdez de Castro, M. J. (2019). Late Middle Pleistocene hominin teeth from Tongzi, southern China. Journal of Human Evolution, 130, 96-108. https://doi.org/10.1016/j.jhevol.2019.03.001
- Liao, W., Xing, S., Li, D., Martinón-Torres, M., Wu, X., Soligo, C., Bermúdez de Castro, M. J., Wang, W., Liu, W. (2019). Mosaic dental morphology in a terminal Pleistocene hominin from Dushan Cave in southern China. Science Reports, 9, 2347. https://doi.org/10.1038/s41598-019-38818-x
- Xing, S., Sun, C., Martinón-Torres, M., Bermúdez de Castro, M. J., Han, F., Zhang, Y., Liu, W. (2016). Hominin teeth from the Middle Pleistocene site of Yiyuan, Eastern China. Journal of Human Evolution, 95, 33-54. https://doi.org/10.1016/j.jhevol.2016.03.004
- Bae, J. C., Wang, W., Zhao, J., Huang, S., Tian, F., Shen, G. (2014). Modern human teeth from Late Pleistocene Luna Cave (Guangxi, China). Quaternary International, 354, 169-183. https://doi.org/10.1016/j.quaint.2014.06.051
- Pan, L., Dumoncel, J., Mazurier, A., Zanolli, C. (2020). Hominin diversity in East Asia during the Middle Pleistocene: A premolar endostructural perspective. Journal of Human Evolution, 148, 102888. https://doi.org/10.1016/j.jhevol.2020.102888
- Pan, L., Dumoncel, J., Mazurier, A., Zanoli, C. (2019). Structural analysis of premolar roots in Middle Pleistocene hominins from China. Journal of Human Evolution, 136, 102669. https://doi.org/10.1016/j.jhevol.2019.102669
- Curnoe, D., Ji, X., Liu, W., Bao, Z., Taçon, C. S. P., Ren, L. (2015). A Hominin Femur with Archaic Affinities from the Late Pleistocene of Southwest China. PLoS ONE, https://doi.org/10.1371/journal.pone.0143332
- Wei, P., Weng, Z., Carlson, J. K., Cao, B., Jin, L., Liu, W. (2021). Late Pleistocene partial femora from Maomaodong, southwestern China. Journal of Human Evolution, 155, 102977. https://doi.org/10.1016/j.jhevol.2021.102977
- “Dali”. PeterBrown-Anthropology (ND). https://www.peterbrown-palaeoanthropology.net/dali.html
- Athreya, S., Wu, X. (2017). A multivariate assessment of the Dali hominin cranium from China: Morphological affinities and implications for Pleistocene evolution in East Asia. American Journal of Biological Anthropology, 164(4): 679-701. https://doi.org/10.1002/ajpa.23305
- Wu, X., Athreya, S. (2012). A description of the geological context, discrete traits, and linear morphometrics of the Middle Pleistocene hominin from Dali, Shaanxi Province, China. American Journal of Biological Anthropology. 150(1): 141-157. https://doi.org/10.1002/ajpa.22188
- “Jinniushan”. PeterBrown-Anthropology (ND). https://www.peterbrown-palaeoanthropology.net/jinniush.html
- “Jinniushan Man Skull (replica)”. National Museum of China, 08-27-21. https://en.chnmuseum.cn/collections_577/collection_highlights_608/archaeological_discoveries_609/202109/t20210927_251612.html
- Rosenberg, R. K., Zuné, L, Ruff, B. C. (2006). Body size, body proportions, and encephalization in a Middle Pleistocene archaic human from northern China. PNAS, 103(10): 3552-3556. https://doi.org/10.1073/pnas.0508681103
- Yang, S., Yue, J., Zhou, X., Storozum, M., Huan, F., Deng, C., Petraglia, D. M. (2020). Hominin site distributions and behaviours across the Mid-Pleistocene climate transition in China. Quaternary Science Reviews, 248, 106614. https://doi.org/10.1016/j.quascirev.2020.106614
- Lu, Z., Meldrum, J. D., Huang, Y., He, J., Sarmiento, E. E. (2011). The Jinniushan hominin pedal skeleton from the late Middle Pleistocene of China. HOMO, 62(6): 389-401. https://doi.org/10.1016/j.jchb.2011.08.008
- “Maba”. PeterBrown-Anthropology (ND). https://www.peterbrown-palaeoanthropology.net/maba.html
- Wu, X., Bruner, E. (2016). The endocranial anatomy of maba 1. American Journal of Biological Anthropology. 160(4): 633-643. https://doi.org/10.1002/ajpa.22974
- Xiao, D., Bae, J. C., Shen, G., Delsin, E., Jin, H. H. J., Webb, M. N., Qiu, L. (2014). Metric and geometric morphometric analysis of new hominin fossils from Maba (Guangdong, China). Journal of Human Evolution, 74, 1-20. https://doi.org/10.1016/j.jhevol.2014.04.003
- Bower, Bruce. “‘Dragon Man’ skull may help oust Neanderthals as our closest relative”. Science News, 06-25-21. https://www.sciencenews.org/article/dragon-man-skull-homo-longi-human-evolution-neandertals
- Wu, X., Pei, S., Cai, Y., Liu, W. (2019). Archaic human remains from Hualongdong, China, and Middle Pleistocene human continuity and variation. PNAS, 116(20): 9820-9824. https://doi.org/10.1073/pnas.1902396116
- Wu, X., Pei, S., Cai, Y., Tong, H., Xing, S., Jashashvili, Carlson, J. K., Liu, W. (2021). Morphological description and evolutionary significance of 300 ka hominin facial bones from Hualongdong, China. Journal of Human Evolution, 161, 103052. https://doi.org/10.1016/j.jhevol.2021.103052
- Chang, C., Kaifu, Y., Takai, M., Kono, T. R., Grün, R., Matsu’ura, S., Kinsley, L., Lin, L. (2015). The first archaic Homo from Taiwan. NaturePorfolio, 27(6): 6037. 10.1038/ncomms7037
- Wu, X., Trinkaus, E. (2014). The Xujiayao 14 Mandibular Ramus and Pleistocene Homo Mandibular Variation. Comptes, Rendus, Palevol, 13(4): 333-341.
- https://doi.org/10.1016/j.crpv.2013.10.002
- Wu, X., S. Pei, Cai, Y., Tong, H., Zhang, Z., Yan, Y., Xing, S., Martinón-Torres, M., Bermúdez de Castro, M. J., Liu, W. (2023). Morphological and morphometric analyses of a late Middle Pleistocene hominin mandible from Hualongdong, China. Journal of Human Evolution, 182, 103411. https://doi.org/10.1016/j.jhevol.2023.103411
- Sáez, Roberto. “A new sister group of the sapiens lineage: on the skulls of Hualongdong and Harbin”. Nutcracker Man, 10-25-21. https://nutcrackerman.com/2021/10/25/nuevo-grupo-hermano-linaje-sapiens-craneos-hualongdong-harbin/
- Bae, J. C. (2010). The late Middle Pleistocene hominin fossil record of eastern Asia: Synthesis and review. American Journal of Biological Anthropology, 143(S51): 75-93. https://doi.org/10.1002/ajpa.21442
- Liu, W., Athreya, S., Xing, S., Wu, X. (2022). Hominin evolution and diversity: a comparison of earlier-Middle and later-Middle Pleistocene hominin fossil variation in China. Philosophical Transactions of the Royal Society B, 377(1848). https://doi.org/10.1098/rstb.2021.0040
Are We The Only Ones to Bury Our Dead? What if we aren’t?

In the realm of human existence, death is an inescapable truth, leaving us to contemplate the fate of our physical forms. Burial, a prevalent practice among humans, serves as a means of honoring the deceased and facilitating the grieving process. However, the question arises: Are we the sole species that engages in the burial of our dead?

Archaeological evidence suggests otherwise. Researchers have uncovered remnants indicating that other hominin species, such as Neanderthals and Denisovans, also practiced burial. This discovery challenges the notion that burial is exclusively a human behavior, underscoring its deep-rooted evolutionary significance.
The reasons behind the practice of burial are multifaceted. Primarily, it offers a ritualized approach to mourning, allowing individuals to process the loss of a loved one. It also reflects a profound respect for the deceased, honoring their memory and acknowledging their contributions to the collective human experience.
Yet, burial may have served an additional purpose. Some scientists propose that our ancestors employed burial as a means of survival. By interring their dead, they could potentially mitigate the spread of diseases and deter predators, thus enhancing the safety and well-being of their communities.
Regardless of the exact motivations, the practice of burial has been an enduring feature of human history. It transcends cultural and geographical boundaries, uniting us in a shared ritual that acknowledges the finite nature of life and the enduring bonds we forge with others.
The realization that other hominin species also practiced burial holds profound implications. It challenges the conventional view that burial is a uniquely human behavior, suggesting instead that it is a deeply ingrained aspect of our evolutionary heritage. This finding raises intriguing questions about the cognitive abilities of our hominin ancestors.
To engage in burial, they must have possessed an understanding of death and mortality, as well as the capacity for forward planning and cooperative action. These findings challenge previous assumptions about the intellectual capabilities of other hominin species, indicating a level of complexity and sophistication that was previously underestimated.
Ultimately, the practice of burial serves as a potent reminder of our shared humanity. It transcends species and connects us to our evolutionary past, underscoring the intricate web of life and the profound interconnectedness of all living beings. Through burial, we honor the memories of those who have come before us, acknowledging their contributions to the rich tapestry of human history and acknowledging the enduring legacy they leave behind.
Hominin Homes

Did other species of humans, in addition to Homo sapiens, have shelters or some other form of artificial structure to make life more comfortable? If so, what can this tell us about different human species’ adaptability and suitability to cold climates?
One possible way to recognize campsites is through the presence of fire. A new technique is helping researchers shed light on the presence of fire at sites where tools and other hominin activity have been found. This technique records any site’s level and molecular density of “polycyclic aromatic hydrocarbons” (or PAHs for short).
PAHs are left behind after a fire, whether that be natural, such as a forest fire, or after a campfire started by humans. The origin of a fire gives rise to a stark difference between the PAHs recorded at the site, both in the structure and frequency of PAHs recorded. Controlled fires by humans leave a much more significant proportion of PAHs with a greater molecular weight than that of forest fires, which produce more substantial numbers of lower molecular weight PAHs.
Lusakert Cave, a Middle Paleolithic site in Armenia known for its Levallois flake tools, is associated with Neanderthals as they were living in the area at the time and were utilizing the Levallois flake tool industry. No specifically modern human tools are known from the site. Researchers working there found many PAHs with a high molecular weight, pointing to hominins’ use of controlled fire as the cause. To be sure, the researchers combined this with what is known about the vegetation and climate of the area at the time the cave was inhabited, some 40-60 thousand years ago. The dating of the relevant layers allowed them to understand when hominins were making a fire in the cave relating to the climatic conditions. They found that fires were produced in the cave even when wildfires would have been reduced in number.
While fire would undoubtedly have been significant during the coldest periods of the Pleistocene epoch (i.e., the Ice Ages), there is evidence of early human species inhabiting temperate to cold regions without the use of fire quite early on. Currently, the first fossil site to illustrate this is Dmanisi, where winter temperatures averaged 2.7ºC (36.86ºF). While by no means cold according to the standards of other periods, for lightly built hominins to inhabit such regions without fire and even quite possibly without clothing shows a level of adaptability within their physical makeup. Later sites such as Happisburgh (pronounced Haysborough) show a similar picture of hominins adapting to cold climates without fire use, despite winters averaging at least 0ºC (32ºF) to -3ºC (26.6ºF).
Migrations as seasons changed could have allowed these early pioneers to avoid the worst of the cold, while staying away from such regions entirely during particularly frigid periods could also have been incorporated into the behavioral repertoire, like that by the hominins at times inhabiting the Nihewan Basin in China. Unfortunately, due to the incompleteness of the fossil record and the vast swathes of time that have passed, it may be tough to notice such strategies. Where examples of early humans that have been present in colder regions do exist, behaviors such as clothing and shelter cannot be exclusively ruled out.
Luckily, archaeological records show that shelters and other structures were designed for comfort. These include several open-air sites in addition to cave sites. These structures vary significantly from site to site in how they are interpreted. For instance, the existence of a collection of large cobbles and boulders 5m (16 and half feet) in length by 1.5m (4 feet and 11 inches) to 1.8m (5 feet 11 inches) in width at the site of Soleihac does appear to resemble a medium-sized dwelling. The researchers working on this site note this and go so far as to point out that water would not have been responsible for the accumulation of the pile of stones. This is because they could not find evidence whatsoever of things such as erosion channels, sheet flow, or even leaching. The researchers also note the presence of a Palaeoloxodon antiquus (straight-tusked elephant) tusk, which, in their words, “was wedged between the rocky blocks.” Additionally, the researchers found a tooth of the same species to be split in two, with one part recovered from the top of the stone pile and the other 0.6m (2ft) below it at the bottom. The researchers describe both parts as fitting together perfectly and cleanly, thus indicating that the pile of stones could have been accumulated during one event.
However, even a conclusion based on this evidence may need to be more solid. Hypothetically, the tooth could have washed in or fallen from a ledge above the site. A high fall could break the tooth, with one part remaining at the top of the pile where it landed, while the other part fell into a crack between the boulders and ended up at the bottom. The pile shifting due to disturbances by later animals, humans, and plants cannot be discounted.
Either way, the Soleihac site highlights an exciting hypothesis and could contradict what was said a few paragraphs above. The site has been dated to 800,000 years ago, which is only about 100,000 years later than the time that the Happisburgh footprints were made. If humans were making shelters some 800,000 years ago further south and in warmer periods, then it would be feasible that they would also be using them in colder regions at colder times.
A real possibility exists that there would have been many shelters in the archaeological record. However, the archaeological record itself may be biased against shelters. For instance, building functioning shelters that keep out the wind and rain would be quite a taxing task to complete daily. One way this can be solved is to stay many nights at any site and have the group’s fittest, most vital, and most complex members travel and hunt or scavenge far from the shelter. They would return before nightfall or when a large hunting party was sent out after several days. In this sense, we could think of the shelter as a staging camp, somewhere the group can return to and meet up before dispersing again. Terra Amata, Melka Kunture, Olorgesailie, Latamne, Bilzingsleben, and Ariendorf 1 have shelters consistent with this model. Where there are caves, there would not be the need to build shelters. However, such sites might still include the construction of windbreaks around the entrance, such as at Le Lazaret and Tor Faraj. This starkly contrasts the many small open-air sites representing more temporary hunting camps.
In conclusion, some evidence supports the argument that other species of humans did use shelters, as outlined above. What can this tell us about different human species’ adaptability and suitability to cold climates? On the one hand, one group of early humans may cope well with freezing conditions thanks to a suite of physiological characteristics, such as higher metabolism, more body hair, and a more significant amount of subcutaneous fat, none of which would show up in the fossil record. On the other hand, a different group devoid of these traits may use ingenuity to adapt its behavior to cope with the climate by building shelters.
Sources:
Ashton, Nick, Simon G. Lewis, Isabelle De Groote, Sarah M. Duffy, Martin Bates, Richard Bates, Peter Hoare, et al. “Hominin footprints from early Pleistocene deposits at Happisburgh, UK.” PLoS One 9, no. 2 (2014): e88329. https://doi.org/10.1371/journal.pone.0088329
Blain, Hugues-Alexandre, Jordi Agustí, David Lordkipanidze, Lorenzo Rook, and Massimo Delfino. “Paleoclimatic and paleoenvironmental context of the Early Pleistocene hominins from Dmanisi (Georgia, Lesser Caucasus) inferred from the herpetofaunal assemblage.” Quaternary Science Reviews 105 (2014): 136-150. https://doi.org/10.1016/j.quascirev.2014.10.004
Bonifay, Eugène, M-F. Bonifay, R. Panattoni, J-J. Tiercelin, and R. Panatonni. “Soleihac (Blanzac, Haute-Loire), nouveau site préhistorique du début du Pléistocène moyen.” Bulletin de la Société préhistorique française. Études et travaux 73 (1976): 293-304.
Brittingham, A., Hren, M.T., Hartman, G. et al. Geochemical Evidence for the Control of Fire by Middle Palaeolithic Hominins. Sci Rep 9, 15368 (2019). https://doi.org/10.1038/s41598-019-51433-0
Cunliffe, Barry, ed. The Oxford illustrated history of prehistoric Europe. Oxford Illustrated History, 2001.
Henry, Donald. “The palimpsest problem, hearth pattern analysis, and Middle Paleolithic site structure.” Quaternary International 247 (2012): 246-266.
Kolen J: Hominids without homes: on the nature of Middle Palaeolithic settlement in Europe. The Middle Palaeolithic occupation of Europe. Roebroeks W, Gamble C, editors. Leiden: University of Leiden and European Science Foundation;1999; pp.139–175.
Lumley H: Terra Amata Nice Alpes-Maritimes. France Tome I, Paris: CNRS Éditions;2009.
Lumley H, Boone Y: Les structures d’habitat au paléolithique inferiéur. La Préhistoire Française. Lumley H, editor. Paris: CNRS;1976; pp.625–643.
Lumley H, Echassoux A, Bailon S, et al.: Le sol d’occupation Acheuléen de l’unité archéostratigraphique UA 25 de la Grotte du Lazaret, Nice, Alpes-Maritimes. Aix-en-Provence: Edisud;2004
MacDonald, Katharine. “The use of fire and human distribution.” Temperature 4, no. 2 (2017): 153-165. https://doi.org/10.1080/23328940.2017.1284637
Parfitt, Simon A., Nick M. Ashton, Simon G. Lewis, Richard L. Abel, G. Russell Coope, Mike H. Field, Rowena Gale et al. “Early Pleistocene human occupation at the edge of the boreal zone in northwest Europe.” Nature 466, no. 7303 (2010): 229-233. https://doi.org/10.1038/nature09117
Roebroeks, Wil, and Thijs Van Kolfschoten. “The earliest occupation of Europe: a short chronology.” Antiquity 68, no. 260 (1994): 489-503. https://doi.org/10.1017/S0003598X00047001
Shaw, Andrew, Martin Bates, Chantal Conneller, Clive Gamble, Marie-Anne Julien, John McNabb, Matt Pope, and Beccy Scott. “The Archaeology of Persistent Places: The Palaeolithic Case of La Cotte de St Brelade, Jersey.” Antiquity 90, no. 354 (2016): 1437–53. https://doi.org/10.15184/aqy.2016.212.
Villa P: Sols at niveaux d’habitat du paleolithique inferieur en Europe et du Proche Orient. Quaternaria. 1976-1977;19:107–134.
Villa P: Conjoinable pieces and site formation processes. Am. Antiq. 1982;47:276–290.
Yang, Shi‐Xia, Cheng‐Long Deng, Ri‐Xiang Zhu, and Michael D. Petraglia. “The Paleolithic in the Nihewan Basin, China: evolutionary history of an early to Late Pleistocene record in Eastern Asia.” Evolutionary Anthropology: Issues, News, and Reviews 29, no. 3 (2020): 125-142. https://doi.org/10.1002/evan.21813
Yar, Bérengère, and Philippe Dubois. “Les structures d’habitat au Paléolithique inférieur et moyen en France: entre réalité et imaginaire.” Bulletin de la Société préhistorique française (1996): 149-163.
Photo credit: Aedrian on Unsplash https://unsplash.com/@aedrian
The Paleo Post Podcast!
The Paleo Post Podcast is a captivating show that dives into the deep history of humanity. Hosted by your friendly neighborhood paleoanthropologists—Seth Chagi, Genevieve von Petzinger, and Dr. George Nash—this podcast makes science fun and accessible. If you’ve ever been confused by headlines about ancient DNA breakthroughs or mind-blowing cave art discoveries, the Paleo Post crew is here to unravel the mysteries for you.
In each episode, they explore topics related to paleogenetics, cave art, and ancient hominin fossils. From ice highways to reanalyzing climate change, they bring you the latest insights from the world of paleoanthropology. So, whether you’re a seasoned enthusiast or a curious listener, the Paleo Post Podcast promises to keep you informed and engaged in the fascinating world of our ancient past.
Remember, our ancestors were once carving tools, painting on cave walls, and navigating a changing world—just like we do today. Tune in and discover the stories that connect us across time and space.
Listen on Apple Podcats and YouTube today!!!
What Would Louis Leakey Say about the Modern Out of Africa Theory – as told by AI
Sometimes AI can be fun, and hearing things in someone else’s voice can be one of those, especially if it’s how to explain something in a certain way.
I hope you enjoy this as much as I did.

“Ah, if I were Louis Leakey, I’d be quite fascinated by the modern interpretations of the Out of Africa theory. You see, when I began my work, we had but a glimpse into the vast expanse of human history. Now, the theory has evolved and expanded with new evidence.
The modern concept still holds that Homo sapiens originated in Africa and spread across the globe. However, recent studies suggest that this migration was not a single exodus but rather a complex series of dispersals over time¹².
One significant update to the theory is the recognition of interbreeding between modern humans and archaic hominins. It’s now understood that as Homo sapiens ventured out of Africa, they occasionally interbred with local populations like the Neanderthals and Denisovans². This paints a picture of a more interconnected prehistoric world than we once thought.
Furthermore, there’s evidence to suggest that the impact of migrations from Asia to Europe may have been stronger than those from Africa, challenging the notion of a purely African origin for all modern humans¹. Teeth morphology studies, for example, show distinct differences between African and Eurasian specimens, indicating a long period of independent evolution on both continents¹.
In essence, while Africa remains the cradle of modern humanity, the story of our spread and interaction with other hominins is far more intricate than I could have ever imagined in my time. It’s a testament to the ever-evolving nature of science and our understanding of human origins..”
Source: Conversation with Bing, 4/12/2024
(1) Is the Out of Africa Theory Out? | Scientific American. https://www.scientificamerican.com/article/is-the-out-of-africa-theory-out/.
(2) Out-of-Africa versus the multiregional hypothesis – Nature. https://www.nature.com/scitable/content/out-of-africa-versus-the-multiregional-hypothesis-6391/.
(3) Modern Humans Came Out of Africa, “Definitive” Study Says. https://www.nationalgeographic.com/history/article/modern-humans-came-out-of-africa-definitive-study-says.
(4) Out of Africa Theory in Evolution | Anthroholic. https://anthroholic.com/out-of-africa-theory.
(5) What is the ‘Out of Africa’ theory? – History Skills. https://www.historyskills.com/classroom/year-7/year-7-human-migration-reading/.
(6) undefined. http://bharatkalyan97.blogspot.in/2013/04/defining-meluhha-demise-of-dravidian.html.
(7) Getty Images. https://www.gettyimages.com/detail/news-photo/british-anthropologist-dr-louis-s-b-leakey-sits-at-a-table-news-photo/3233313.
